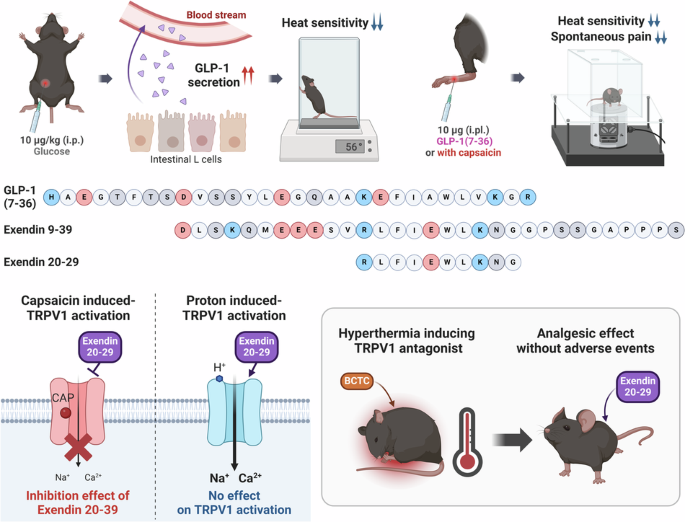
Glp-1 and its derived peptides mediate pain relief through direct trpv1 inhibition without affecting thermoregulation
- Select a language for the TTS:
- UK English Female
- UK English Male
- US English Female
- US English Male
- Australian Female
- Australian Male
- Language selected: (auto detect) - EN
Play all audios:

ABSTRACT Hormonal regulation during food ingestion and its association with pain prompted the investigation of the impact of glucagon-like peptide-1 (GLP-1) on transient receptor potential
vanilloid 1 (TRPV1). Both endogenous and synthetic GLP-1, as well as a GLP-1R antagonist, exendin 9–39, reduced heat sensitivity in naïve mice. GLP-1-derived peptides (liraglutide,
exendin-4, and exendin 9–39) effectively inhibited capsaicin (CAP)-induced currents and calcium responses in cultured sensory neurons and TRPV1-expressing cell lines. Notably, exendin 9–39
alleviated CAP-induced acute pain, as well as chronic pain induced by complete Freund’s adjuvant (CFA) and spared nerve injury (SNI), in mice without causing hyperthermia associated with
other TRPV1 inhibitors. Electrophysiological analyses revealed that exendin 9–39 binds to the extracellular side of TRPV1, functioning as a noncompetitive inhibitor of CAP. Exendin 9–39 did
not affect proton-induced TRPV1 activation, suggesting its selective antagonism. Among the exendin 9–39 fragments, exendin 20–29 specifically binds to TRPV1, alleviating pain in both acute
and chronic pain models without interfering with GLP-1R function. Our study revealed a novel role for GLP-1 and its derivatives in pain relief, suggesting exendin 20–29 as a promising
therapeutic candidate. SIMILAR CONTENT BEING VIEWED BY OTHERS INHIBITING HV1 CHANNEL IN PERIPHERAL SENSORY NEURONS ATTENUATES CHRONIC INFLAMMATORY PAIN AND OPIOID SIDE EFFECTS Article 03
February 2022 STRUCTURAL BASIS OF TRPV1 MODULATION BY ENDOGENOUS BIOACTIVE LIPIDS Article Open access 02 May 2024 STRUCTURE-GUIDED PEPTIDE ENGINEERING OF A POSITIVE ALLOSTERIC MODULATOR
TARGETING THE OUTER PORE OF TRPV1 FOR LONG-LASTING ANALGESIA Article Open access 03 January 2023 INTRODUCTION Individuals experiencing pain often report overeating calorie-dense, high-sugar,
and high-fat foods1 as a coping mechanism2. This phenomenon, termed “ingestion analgesia,” has also been observed in animals3,4,5,6. Noxious heat-evoked withdrawal behaviors in rats are
suppressed during self-initiated chocolate eating and the ingestion of sucrose7. Although much research has focused on mechanisms within the central nervous system to explain this pain
suppression8, we hypothesize that additional mechanisms occurring in the peripheral nervous system are based on hormonal regulation during digestion. Upon food ingestion and its subsequent
entry into the small intestine, intestinal L-cells undergo posttranslational processing of the proglucagon gene, leading to the production of glucagon-like peptide-1 (GLP-1), a potent
incretin peptide hormone9. GLP-1 plays an important role in glucose homeostasis and is secreted into the hepatic portal system in response to elevated glucose levels from food intake10,
stimulating insulin synthesis and release from the pancreas11. The short half-life of native GLP-1, typically only 1–2 min due to rapid degradation, has facilitated the development of
structurally modified GLP-1 analogs with longer half-lives, such as liraglutide, exendin-4, and dulaglutide, which exhibit 97%, 53%, and 90% sequence homology, respectively12. Regarding pain
modulation, various studies have reported that GLP-1 analogs exert antinociceptive effects on the central nervous system. This modulation occurs by regulating the spinal dorsal horn
microglial pathway through GLP-1R activation, leading to alterations in spinal excitatory synaptic transmission under neuropathic pain conditions13,14,15,16,17,18. However, the effects of
GLP-1 on the peripheral nervous system and its relationship with pain modulation remain largely unexplored. The transient receptor potential vanilloid 1 (TRPV1) channel plays a crucial role
in heat pain perception19. TRPV1, known for its thermosensitive and polymodal nociceptor properties, is expressed in sensory neurons and is activated by stimuli such as capsaicin (CAP) and
protons20. In chronic pain states, TRPV1 channels are upregulated in nociceptive neurons, which lowers stimulation thresholds and increases pain perception, as reported in hyperalgesia or
allodynia21. Inhibiting TRPV1 has proven effective in mitigating pain in diverse neuropathic pain models, thereby garnering interest from numerous pharmaceutical companies22,23. However, the
challenge with current TRPV1 antagonists lies in their associated adverse effects, such as hyperthermia24,25,26,27,28,29,30,31 and hypothermia32,33,34, caused by failures in
thermoregulation. This issue has been attributed to the mode of action of these antagonists: those blocking CAP-, proton-, and heat-induced TRPV1 activation result in hyperthermia, whereas
those sparing proton-induced activation do not lead to hyperthermia33,35. Conversely, the potentiation of proton-induced TRPV1 activation leads to hypothermia32. In light of these
challenges, in this study, we assess the impact of GLP-1 and its derivatives on pain behavior and their influence on the peripheral nervous system. Preliminary findings indicate that GLP-1
harbors a key sequence that directly binds to and inhibits the activation of TRPV1 channels in sensory neurons. Importantly, we also demonstrate that GLP-1 and its derivatives directly
antagonize TRPV1 channels in a mode-selective manner that can offer pain relief without the adverse thermoregulatory side effects commonly associated with current TRPV1 inhibitors. MATERIALS
AND METHODS CHEMICALS CAP and N-(4-tertiarybutylphenyl)-4-(3-cholorphyridin-2-yl) tetrahydropyrazine-1(2H)-carboxamide (BCTC) were purchased from Sigma‒Aldrich (St. Louis, MO), and the
stock solutions were prepared with 99.5% ethanol and dimethyl sulfoxide, respectively. AITC and CFA were purchased from Sigma‒Aldrich. GLP-1(7-36), liraglutide, exendin-4, exendin 9–39,
exendin 20–29, His-tagged exendin 9–39, and FITC-tagged exendin 9–39 were purchased from Anygen Corp. (Gwangju, South Korea). The stock solutions were prepared according to the supplier’s
recommendation for each peptide and were stored at −20 °C. The exendin 20–29 alanine scanning library was purchased from GenScript (Piscataway, NJ). ANIMALS Adult wild-type male C57BL/6 N
mice were purchased from Orient Bio (Sungnam, South Korea). The mice were housed at a constant temperature and humidity under a 12-h/12-h light‒dark cycle with free access to standard food
and water for at least 1 week prior to the beginning of the experimental procedures. All animal experiments were approved by the Institutional Animal Care and Use Committee of the College of
Medicine at Gachon University (approval number: LCDI-2020-0135). CELL PREPARATION AND TRANSIENT TRANSFECTION The dorsal root ganglia (DRGs) were aseptically removed from 5–8-week-old mice
and incubated with collagenase A (0.2 mg/mL; Roche, Basel, Switzerland)/dispase-II (2.4 units/mL; Roche) at 37 °C for 90 min. The cells were mechanically dissociated via gentle pipetting and
placed on glass coverslips coated with poly-D-lysine. DRG cells were then grown in neurobasal medium supplemented with 10% fetal bovine serum (FBS; Gibco, Waltham, MA), 2% B27 supplement
(Invitrogen, Carlsbad, CA), and 1% penicillin/streptomycin for 24 h before the experiments. Human TRPV1 (hTRPV1)-expressing CHO K1 cells were cultured in Dulbecco’s modified Eagle’s medium
(DMEM) supplemented with 10% FBS, 1% penicillin/streptomycin, and 800 µg/mL geneticin in an incubator with 5% CO2 at 37 °C. HEK293 and HEK293T cells were also cultured in the same DMEM but
without geneticin. Rat TRPV1 plasmids were labeled with green fluorescent protein for subsequent calcium imaging. cDNA constructs of rat TRPV1 were transiently transfected into cells via
Lipofectamine 2000 (Invitrogen). Ca2+ imaging was performed one day after transient transfection. PATCH-CLAMP RECORDINGS Whole-cell voltage-clamp and cell-attached patch-clamp recordings
were conducted at room temperature to measure capsaicin (CAP)- or proton-induced currents in dissociated DRG neurons and hTRPV1-expressing CHO K1 cells, respectively. An EPC10 amplifier
(HEKA, Stuttgart, Germany) was utilized for these recordings. Patch pipettes, prepared with a micropipette puller (Narishige, Tokyo, Japan), had resistances of 4–5 MΩ for whole-cell
recordings and 6–8 MΩ for cell-attached recordings. The recording chamber, with a volume of 500 µL, was continuously superfused at a rate of 1–2 mL/min. CAP- or low pH-induced currents were
recorded at a holding potential of −60 mV. Series resistance was compensated ( > 80%), and leak subtraction was performed. The data acquired were low-pass filtered at 2 kHz, sampled at 10
kHz, and analyzed via PatchMaster and FitMaster (HEKA) software. The internal solution for whole-cell and cell-attached recordings consisted of 126 mM K-gluconate, 10 mM NaCl, 1 mM MgCl2,
10 mM EGTA, 10 mM HEPES, 2 mM NaATP, and 0.1 mM Na2GTP, adjusted to pH 7.3 with KOH, with an osmolarity of 295–300 mOsm. An external solution of 140 mM NaCl, 5 mM KCl, 1 mM MgCl2, 2 mM EGTA,
10 mM HEPES, and 10 mM glucose, adjusted to pH 7.4 with NaOH, osmolarity 300–310 mOsm, was used for both techniques. Inside-out recordings from hTRPV1-expressing CHO K1 cells were conducted
using a pipette resistance of 8–9 MΩ. The internal solution for the inside-out recordings matched that used for the whole-cell and cell-attached methods, whereas the external solution
(intracellular side) differed slightly, containing 2 mM CaCl2 instead of EGTA. The open probability and average single-channel opening and closing times for the inside-out recordings were
analyzed via a 50% threshold criterion, with all events double-checked before analysis. CA2+ IMAGING IN DISSOCIATED DRG NEURONS AND HTRPV1-CHO K1 CELLS At room temperature, Ca2+ imaging was
conducted in mouse DRG, HEK293T, and hTRPV1-expressing CHO K1 cells. Cells on poly-D-lysine-coated coverslips were loaded with 2 µM Fura-2 AM (Thermo Fisher Scientific, Waltham, MA) at 37 °C
for 40 min in DMEM. The cells were then rinsed three times with the medium and incubated for 30 min, after which they were placed on the stage of an inverted microscope (BX51W1; Olympus,
Tokyo, Japan) and continuously superfused at a flow speed of 1 mL/min with a bath solution containing 140 mM NaCl, 5 mM KCl, 1 mM CaCl2, 2 mM MgCl2, 10 mM HEPES, and 10 mM glucose, adjusted
to pH 7.4 with NaOH. Using illumination with a 175-W xenon arc lamp, excitation wavelengths (340/380 nm) were selected via a Lambda DG-4 monochromator wavelength changer (Shutter Instrument,
Novato, CA). The fluorescence 340/380 ratio was measured via digital video microfluorometry with an intensified camera (OptiMOS; QImaging, Surrey, Canada) coupled to the microscope. The
data were analyzed via SlideBook 6 (Intelligent Imaging Innovations, Denver, CO). At the end of the experiment, the cells were identified on the basis of their response to high
concentrations of KCl. PULL-DOWN ASSAY AND IMMUNOBLOTTING Interactions between exendin 9–39 and TRPV1 were examined via His-mediated pull-down assays using a modified protocol from the
Pierce Pull-down PolyHis Protein:Protein Interaction Kit (Thermo Fisher Scientific). His-tagged exendin 9–39 was bound to HisPur Cobalt Resin as the bait protein and incubated with lysates
of hTRPV1-CHO K1 or native CHO K1 cells overnight at 4 °C. After washing five times with lysis buffer, the complexes were mixed with lysis buffer containing 290 mM imidazole, and the bound
proteins were eluted by boiling in 5× sodium dodecyl sulfate (SDS) loading buffer for 5 min. The products were then separated via SDS‒polyacrylamide gel electrophoresis, transferred onto
nitrocellulose membranes, and blotted with an anti-TRPV1 antibody (#ACC‒030; Alomone Labs, Jerusalem, Israel). The membrane was washed with TBST and incubated for 1 h with the secondary
antibody, anti-rabbit Ig-HRP (1:10,000, 9910; Cell Signaling Technology, Danvers, MA). After washing with TBST, the immune complexes were detected via chemiluminescence (Beyotime, Shanghai,
China) using a Pierce western blotting kit (Thermo Fisher Scientific). Quantitative densitometric analysis was performed via a UVP BioSpectrum multispectral imaging system (Image Quant LAS
4000; GE Healthcare, Chicago, IL). ELISA The mice were anesthetized using isoflurane, and blood samples were collected at specified time points. Blood was drawn before (0 min) and after the
intraperitoneal administration of glucose (2.0 g/kg body weight) at intervals of 2, 6, 10, 30, 60, and 120 min (_n_ = 3–4 per time point). Serum was obtained by centrifuging the collected
whole blood at 3000 ×g for 15 min at 4 °C. The supernatant was then transferred to a fresh tube, and total GLP-1 levels were measured via an ELISA kit (EZGLP1T-36K, Millipore, Tokyo, Japan)
following the manufacturer’s instructions. CONFOCAL FLUORESCENCE IMAGING Approximately 1 × 105 mL−1 of hTRPV1-expressing CHO K1 cells or naïve CHO K1 cells were seeded into confocal dishes
and cultured overnight for cell adherence. The cells were washed with phosphate-buffered saline (PBS) and fixed with 2% paraformaldehyde for 10 min at 25 °C. After being washed with PBS, the
cells were permeabilized with 0.1% Triton X-100 for 5 min at room temperature and blocked with 3% bovine serum albumin with glycine for 30 min. The cells were incubated with a primary
antibody against TRPV1 (1:250; #ACC-030, Alomone Labs) for 1 h at room temperature. After washing with PBS three times, an Alexa Fluor 594-conjugated secondary antibody (1:400; Invitrogen)
and Hoechst 33342 (Thermo Fisher Scientific) were added for another hour of incubation at 4 °C. The cells were washed with PBS and incubated with FITC-tagged exendin 9–39 dissolved in cold
PBS at a concentration of 10 μM for 30 min at 4 °C. Finally, after washing with cold PBS, images were acquired via a confocal laser-scanning microscope with a 100× oil-immersion objective
(LSM 700; Carl Zeiss, Oberkochen, Germany). BEHAVIORAL TESTS IN MICE Baseline heat sensitivity was assessed to evaluate the systemic effects of glucose or GLP-1(7–36) administration via a
hot plate apparatus (Ugo Basile, Italy). The paw withdrawal latency (PWL) to heat was measured after either intraperitoneal or oral gavage administration of glucose (2.0 g/kg body weight) in
a volume of 200 μL or the same volume of vehicle (0.9% saline) (_n_ = 5 per group). To test the local effects of GLP-1(7–36) and exendin 9–39 on heat sensitivity, PWL was measured via the
Hargreaves radiant heat apparatus following intraplantar administration of 10 μg of GLP-1(7–36) or vehicle (0.9% saline) (_n_ = 5 per group) in a volume of 10 μL. To evaluate nociceptive
behavior, the time spent licking the paw, the paw withdrawal threshold (PWT) to mechanical stimuli, and the PWL to heat were measured as described previously36. Either vehicle, exendin 9–39,
exendin 20–29, or BCTC was first injected via the intraplantar route, and 30 min later, an additional injection of exendin 9–39, exendin 20–29, or BCTC in combination with CAP was
administered. Licking time was recorded for 5 min after CAP injection in each group. Mechanical allodynia and thermal hyperalgesia were evaluated in separate experiments in a time-dependent
manner. Mechanical allodynia was assessed via von Frey filaments (NC12775-99; North Coast Medical, CA). The 50% PWT was calculated via the updown method. Thermal hyperalgesia was assessed by
recording the PWL via the Hargreaves radiant heat apparatus (IITC Life Sciences, Woodland Hills, CA). A cutoff value of 20 s was used to prevent tissue damage. For rectal temperature
recording, the rectal temperature was measured with a digital thermometer (Therma‐1; ETI, West Sussex, UK) by inserting a corn oil-soaked flexible bead probe into the rectum after the
intraperitoneal administration of 200 μL of vehicle, exendin 9–39 (50 µg/kg), or BCTC 5 mg/kg in wild-type mice (_n_ = 6). The acute intraperitoneal glucose tolerance test (IPGTT) was
performed in 8-week-old mice. Baseline blood glucose levels were measured, and the wild-type mice were intraperitoneally injected with 200 μL of vehicle, exendin 20–29 (10 μg/kg), or
exendin-4 (10 μg/kg) (_n_ = 6). After 15 min, the mice were challenged with glucose (2.0 g/kg body weight). Blood glucose levels were measured via an Accu-Chek Performa glucometer (Roche,
Mannheim, Germany). CFA-INDUCED PAIN Under temporal anesthesia with 3% isoflurane, the CFA-induced inflammatory pain model was established via the intraplantar injection of 20 μL of CFA. The
mice were then treated with either vehicle, exendin 9–39, or exendin 20–29 in 20 μL, which were administered on the same paw where the CFA was injected. Heat hyperalgesia and mechanical
allodynia induced by CFA were assessed via the Hargreaves test and the von Frey test, respectively. Paw thickness was measured in millimeters via a CD-15APX digimatic caliper (Mitutoyo
Corporation, Kawasaki, Japan). SNI-INDUCED PAIN Under continuous anesthesia with isoflurane, mice underwent surgical manipulation to expose the left sciatic nerve by separating the muscle
tissue. Upon visualization of the sciatic nerve, the peroneal and tibial nerves were ligated and transected at the lower end of the ligature via silk thread, whereas the sural nerve remained
intact. The surgical site was then sutured, and iodine was applied for debridement. After a recovery period of 14 days postsurgery, the mice received either intraplantar or intraperitoneal
administration (20 or 200 μL, respectively) of vehicle, exendin 9–39, or exendin 20–29. The heat sensitivity of the neuropathic pain model mice was subsequently assessed via the Hargreaves
test. STATISTICAL ANALYSIS Statistical analyses were conducted via GraphPad Prism 8 (GraphPad Software, San Diego, CA). All the data are presented as the mean ± standard error of the mean
(S.E.M.). Differences between groups were compared via two-tailed unpaired _t_ tests for two groups, one-way analysis of variance (ANOVA) followed by Dunnett’s multiple comparison test for
multiple groups, or two-way repeated-measures ANOVA followed by the Bonferroni multiple comparison test for multiple groups and time courses. The statistical significance thresholds were
*_p_ < 0.05, **_p_ < 0.01, ***_p_ < 0.001, and ****_p_ < 0.0001 (likewise indicated with # and †). RESULTS RELEASE OF GLP-1 BY GLUCOSE APPLICATION ALLEVIATES HEAT SENSITIVITY
Given the essential role that GLP-1 plays in regulating glucose levels in the body, we primarily utilized intraperitoneal glucose administration and assessed changes in heat sensitivity in
mice via hot plate tests. The role of endogenous GLP-1 in heat sensitivity was demonstrated by the observation that the intraperitoneal glucose-treated group presented a decrease in heat
sensitivity compared with the vehicle-treated group (Fig. 1a), as the administration of glucose significantly increased blood glucose levels from 1 to 2 h (Supplementary Fig. 1a). To further
explore the relationship between glucose-induced GLP-1 release and heat sensitivity, we examined changes in total GLP-1 levels in the serum following glucose administration (Supplementary
Fig. 1b). The data revealed a significant increase in GLP-1 levels, peaking at 30 min postinjection and subsequently decreasing over time. This peak in GLP-1 concentration correlated with
the observed changes in heat sensitivity, supporting the hypothesis that the systemic release of GLP-1 contributes to the analgesic effects. To confirm whether the reduction in heat
sensitivity after glucose application was due to the systemic release of GLP-1, we intraperitoneally administered 10 µg/kg GLP-1(7–36), one of the two primary biologically active forms of
secreted GLP-1. Similar to the results observed with glucose administration, the hot plate test revealed a reduction in heat sensitivity after 1 h (Fig. 1b, f). Further exploration of GLP-1
administration routes led us to assess the effects of the oral administration of GLP-1(7–36). Remarkably, this method demonstrated a reduction in heat sensitivity that was comparable to, or
even exceeded, that observed with intraperitoneal administration (Fig. 1c). This observation underscores the potential of the oral delivery of GLP-1(7–36) as a feasible method for pain
prevention. Although our study did not directly evaluate the analgesic effects of oral glucose administration, the analgesic outcomes associated with oral GLP-1(7–36) suggest that increased
GLP-1 secretion triggered by oral glucose could similarly influence pain responses. Typically, GLP-1 secretion in response to oral glucose is more pronounced than that in response to other
administration routes37, suggesting a significant opportunity for future investigations to directly examine the analgesic effects of oral glucose through its impact on GLP-1 levels. Upon
discovering the impact of systemic GLP-1 on heat sensitivity, we evaluated its influence on the peripheral nervous system through local intraplantar injection of 10 µg of GLP-1(7–36). The
results of the Hargreaves test demonstrated a reduction in pain sensitivity (Fig. 1d). As changes in heat sensitivity are dependent on TRPV1 activity, we sought to determine whether
intraplantar injection of 10 µg of GLP-1(7–36) modulates spontaneous pain behavior induced by the TRPV1 agonist CAP in mice. We found that intraplantar injection of 10 µg of GLP-1(7–36) for
10 min significantly reduced the pain-like (licking) behavior induced by intraplantar injection of 1.6 µg of CAP (Fig. 1e). Therefore, we hypothesized that GLP-1 and its metabolites affect
peripheral pain regulation through the TRPV1 channel (Fig. 1g). GLP-1 AND ITS ANALOGS MODULATE CAP-INDUCED TRPV1 ACTIVATION IN MOUSE DRG NEURONS To determine whether GLP-1 regulates
peripheral pain signaling by modulating TRPV1 activity in sensory neurons, whole-cell patch-clamp recordings were performed using small-diameter (<25 µm) neurons dissociated from the
mouse DRG. The application of 100 nM CAP elicited inward currents, indicating CAP-responsive DRG neurons, and these responses were abolished by pretreatment with 100 nM GLP-1(7–36) by 52%
(Fig. 2a, f). We further tested TRPV1 channel modulation via Ca2+ imaging (Fig. 2b–e, g). Pretreatment with each examined GLP-1 analogs, including GLP-1(7–36), liraglutide, and exendin-4,
led to significantly reduced Ca2+ responses (by 53.8%, 58.1%, and 72.2%, respectively). These results suggest that GLP-1 analogs play an antagonistic role in regulating TRPV1 function. THE
GLP-1 ANTAGONIST, EXENDIN 9–39, REGULATES NOCICEPTION VIA TRPV1 MODULATION Exendin 9–39, a synthetic peptide, functions as a specific and competitive antagonist of GLP-138. It is derived
from exendin-4 through N-terminal truncation and shares 53% sequence homology with native GLP-139. We initially sought to examine whether exendin 9–39 could reverse the decrease in pain
sensitivity caused by local intraplantar injection. However, the results of the Hargreaves test indicated that the administration of 10 µg of exendin 9–39 decreased heat sensitivity upon
intraplantar injection (Fig. 3a). To determine its effects, we pretreated DRG cultures with 100 nM exendin 9–39. Like GLP-1, both CAP-induced TRPV1 inward currents and Ca2+ responses were
decreased by 85% and 90.6%, respectively (Fig. 3b, c). To determine the direct effect of exendin 9–39 on the time that mice spent licking their hind paws in the CAP-induced spontaneous pain
model, intraplantar administration of exendin 9–39 (doses of 5 and 10 μg) was followed by intraplantar administration of CAP (1.6 μg). Exendin 9–39 dose-dependently reduced the paw licking
time, similar to the effect observed with BCTC (0.5 μg), which was used as the control (Fig. 3d). In addition, the paw withdrawal latency (PWL) in the Hargreaves test was significantly lower
in the vehicle + CAP group than in the vehicle group from 30 to 120 min (Fig. 3f). Similarly, compared with vehicle injection, intraplantar CAP injection significantly reduced the PWT in
the von Frey test from 30 to 120 min compared with that in the vehicle group (Fig. 3f), whereas mechanical allodynia was dose-dependently alleviated in the groups treated with exendin 9–39
(5 or 10 μg) or BCTC (0.5 μg), a widely used TRPV1 inhibitor40. These results suggest that the analgesic effects of exendin 9–39 via TRPV1 are similar to those of BCTC. To determine whether
exendin 9–39 treatment causes hyperthermia, which is a typical adverse effect of TRPV1 inhibitors41, body temperature was measured following the administration of exendin 9–39. Previous
animal studies have shown that the intraperitoneal administration of various TRPV1 antagonists results in hyperthermia42; therefore, in this study, exendin 9–39 was injected
intraperitoneally. As exendin 9–39 had analgesic effects on the nociceptive behavior of mice at a mass 10 times greater than that of BCTC, the same factor (50 mg/kg exendin 9–39 vs. 5 mg/kg
BCTC) was used in this experiment. The rectal temperature in the group treated with exendin 9–39 was similar to that in the vehicle-treated group. In contrast, the rectal temperature in the
BCTC-treated group rapidly increased by 0.7–1.5 °C and returned to baseline values at 12 min postadministration (Fig. 3e). EXENDIN 9–39 ALLEVIATES INFLAMMATORY AND NEUROPATHIC CHRONIC PAIN
BEHAVIORS The analgesic potential of exendin 9–39 was confirmed in an acute pain mouse model, followed by an assessment of its effectiveness in a chronic pain model. Considering the
documented changes in sensitization and upregulation of TRPV1 in the complete Freund’s adjuvant (CFA)-induced inflammatory pain model, CFA was utilized to induce chronic inflammatory pain in
mice43. The effects of exendin 9–39 on heat hyperalgesia and mechanical allodynia were assessed via the Hargreaves test and the von Frey test, respectively (Fig. 4a). In the Hargreaves
test, mice with CFA-induced inflammation displayed delayed recovery from heat hyperalgesia following intraplantar vehicle administration. However, a single intraplantar administration of 5
or 10 μg of exendin 9–39 effectively relieved heat hyperalgesia and accelerated the recovery process (Fig. 4b). Similarly, in the von Frey test, mice injected with the intraplantar vehicle
after CFA induction displayed prolonged mechanical allodynia, which was significantly reversed by a single intraplantar administration of 5 or 10 μg of exendin 9–39 (Fig. 4c). Additionally,
exendin 9–39 administration mitigated paw swelling resulting from CFA induction (Supplementary Fig. 2). Furthermore, peripheral nerve injury leads to neuropathic pain, and the spared nerve
injury (SNI) model in mice induces persistent heat hyperalgesia44. Fourteen days after SNI surgery, compared with those in the sham group, the mice in the SNI group presented heat
hyperalgesia after intraplantar vehicle administration (Fig. 4d). However, SNI-challenged mice administered intraplantar injections of 10 μg of exendin 9–39 presented alleviated heat
hyperalgesia starting at 1 h postadministration, peaking at 3 h, compared with those in the vehicle-administered group (Fig. 4e). Similarly, when the same dose of exendin 9–39 was
administered intraperitoneally to test its systemic analgesic effects, it alleviated heat hyperalgesia starting at 1 h postadministration, peaking at 3 h, compared with the effects of the
intraperitoneal administration of the vehicle (Fig. 4f). These findings suggest that the analgesic effects of exendin 9–39 extend not only to chronic inflammatory pain but also to
neuropathic pain. GLP-1-DERIVED PEPTIDES REGULATE RAT AND HUMAN TRPV1 ACTIVATION INDEPENDENTLY OF GLP-1R EXPRESSION To verify the potential involvement of GLP-1R in the observed inhibitory
effects on TRPV1 channels, we transiently transfected rat TRPV1 into HEK293T cells. GLP-1R expression was not detected in the HEK293T cell line before transfection (Supplementary Fig. 3).
CAP-induced TRPV1 calcium responses were again inhibited by pretreatment with GLP-1 analogs or exendin 9–39 (Fig. 5a–e), indicating that no interaction with GLP-1R was involved. Next, we
estimated and compared the half-maximal inhibitory concentration (IC50) values from the concentration‒response data using Chinese hamster ovary (CHO K1) cells expressing human TRPV1. The
IC50 values for the calcium response of GLP-1, liraglutide, exendin-4, and exendin 9–39 were 178.60, 62.19, 64.49, and 28.18 nM, respectively (Fig. 5f–h). To determine the subtype
selectivity of exendin 9–39, we tested its effect on TRPA1. Exendin 9–39 did not inhibit or potentiate TRPA1 currents induced by 100 μM allyl isothiocyanate (AITC) (Supplementary Fig. 4a–c),
demonstrating that exendin 9–39 is a selective inhibitor of TRPV1. As exendin 9–39 had the lowest IC50 value for inhibiting CAP-activated TRPV1 currents, its molecular mechanism was further
analyzed. EXENDIN 9–39 DIRECTLY BINDS TO THE TRPV1 CHANNEL To assess the direct interaction between exendin 9–39 and the TRPV1 channel, we conducted a protein binding assay using His-tagged
exendin 9–39, which was able to pull down the TRPV1 channel (Fig. 5i) in the CHO K1 cell line stably expressing human TRPV1. In contrast, the naïve CHO K1 cell line showed no TRPV1 channel
binding. We tested the binding of exendin 9–39 to TRPV1 by incubating the two cell lines with fluorescein isothiocyanate (FITC)-labeled exendin 9–39. Immunofluorescence imaging revealed that
FITC-labeled exendin 9–39 was bound to TRPV1 on the cell surface in CHO K1 cells expressing human TRPV1 but not in naïve CHO K1 cells (Fig. 5j). Although exendin 9–39 was tagged with FITC,
it still blocked the CAP-induced inward currents of TRPV1 channels (Supplementary Fig. 5a, b). EXENDIN 9–39 TARGETS THE EXTRACELLULAR SIDE OF THE TRPV1 CHANNEL BUT DOES NOT SHARE THE CAP
BINDING SITE Next, we examined the molecular target of exendin 9–39 on the TRPV1 channel via a patch‒clamp competition assay in CHO K1 cells expressing human TRPV1. BCTC, a highly potent
TRPV1 antagonist that competitively interacts with CAP to bind to TRPV1 channels40, was used for this assay. BCTC (10 nM) was applied in the presence of 10 nM CAP until saturation, and twice
the BCTC concentration (20 nM) was sequentially applied. Although 10 nM BCTC blocked the CAP-induced TRPV1 currents, 20 nM BCTC did not significantly potentiate this TRPV1-inhibiting effect
(Fig. 6a). In contrast, 100 nM exendin 9–39 application following 10 nM BCTC elicited significant further inhibition of CAP-evoked TRPV1 inward currents (Fig. 6b). These results indicate
that exendin 9–39 does not share a binding site with CAP and inhibits TRPV1 activation by interacting noncompetitively with CAP. To validate the TRPV1 binding site, we performed
single-channel recordings from cells expressing human TRPV1 channels. Cell-attached patch recordings revealed that the external bath application of 10 nM CAP and 100 nM exendin 9–39 did not
reduce the frequency of single-channel open events. In contrast, when the pipette solution contained 100 nM exendin 9–39, which allowed the interaction of exendin 9–39 with the extracellular
surface of TRPV1, single-channel opening events were significantly blocked (Fig. 6c–e). We further conducted inside-out patch-clamp recordings to determine whether bath application of CAP
and exendin 9–39 to the intracellular surface of TRPV1 changed single-channel opening events (Fig. 6f–h). The results showed that single-channel openings were blocked by CAP applied to the
intracellular side of the channel only when the pipette solution contained 100 nM exendin 9–39, facilitating the exposure of exendin 9–39 to the extracellular surface of TRPV1. In contrast,
single-channel opening events were not detected when CAP and exendin 9–39 were administered via the external bath, allowing exposure of CAP and exendin 9–39 to the intracellular surface of
TRPV1. Thus, exendin 9–39 binds to the extracellular side of TRPV1 but cannot penetrate the membrane. EXENDIN 9–39 DOES NOT AFFECT TRPV1 ACTIVATION VIA PROTONS, SUGGESTING ITS MODE
SELECTIVITY As the inhibitory effect of exendin 9–39 on CAP-induced activation of TRPV1 was confirmed, its effect on proton-induced activation of TRPV1 was also tested via patch-clamp
recordings and calcium imaging. BCTC is also a potent antagonist of TRPV1 activated by protons45; therefore, it was used as a control antagonist. Unlike exendin 9–39 during CAP-induced TRPV1
activation, 1 μM exendin 9–39 neither reduced nor potentiated low pH-induced inward currents (Fig. 7a, b) and calcium influx (Fig. 7c, d) in hTRPV1-expressing CHO K1 cells, whereas 1 μM
BCTC completely blocked these currents. The exendin 9–39 concentration used in this experiment was approximately 30 times higher than its IC50 value for CAP-induced TRPV1 activation,
indicating the mode-selective characteristics of exendin 9–39. These findings suggest that exendin 9–39 may be less likely to cause adverse effects associated with thermoregulation, such as
hyperthermia or hypothermia, as observed with previous TRPV1 antagonists. SMALL PEPTIDES DERIVED FROM EXENDIN 9–39 INHIBIT TRPV1 ACTIVATION AND ATTENUATE CAP-INDUCED SPONTANEOUS PAIN IN MICE
To verify the lack of GLP-1R involvement in the observed effects of exendin 9–39, we roughly fragmented exendin 9–39 into three small peptides, i.e., exendin 9–29, 14–32, and 20–39 (Fig.
8a), all of which reduced CAP-induced inward currents (Fig. 8b) and calcium responses (Fig. 8c). Therefore, we assumed that the overlapping peptide sequence exendin 20–29 plays a key role in
the inhibition of CAP-induced TRPV1 activation. As hypothesized, exendin 20–29 showed similar inhibitory effects (Fig. 8b, c). Further structural analysis via the program AlphaFold46 was
conducted to visualize and better comprehend the interactions of the exendin 20–29 sequence with TRPV1 within the broader context of GLP-1 and its derivatives (Supplementary Fig. 6). This
analysis highlighted the unique binding potential of exendin 20–29, suggesting its efficacy as a targeted TRPV1 blocker derived from exendin 9–39. The analgesic effects of exendin 20–29 were
then examined in the same manner as those of exendin 9–39. Intraplantar administration of exendin 20–29 (20 μg) also reduced the paw licking time (Fig. 8d). Compared with intraplantar
administration of CAP alone (1.6 μg), additional intraplantar injection of exendin 20–29 (20 μg) significantly alleviated pain from 60 to 120 min (Fig. 8e), as manifested by increased PWL in
the Hargreaves test. This alleviation was also observed in the group with additional BCTC (0.5 μg) treatment, and no significant differences between the 20 μg exendin 20–29 + CAP and 0.5 μg
BCTC + CAP groups were found. Similarly, intraplantar exendin 20–29 (20 μg) injection significantly increased the PWT in the von Frey test from 30 to 90 min compared with CAP (1.6 μg)
administration alone (Fig. 8e). However, no significant differences were found between the 20 μg exendin 20–29 + CAP and 0.5 μg BCTC + CAP groups. We also determined whether exendin 20–29
modulates proton-induced TRPV1 activation via patch-clamp recordings and calcium imaging. When 1 μM exendin 20–29 was applied, no significant effect was found, in contrast to the 92%
inhibition elicited by 1 μM BCTC (Supplementary Fig. 7a–c). Collectively, these data reveal that exendin 20–29 acts as a key sequence in inducing analgesia in CAP-elicited nociceptive
behaviors in mice without causing hyperthermia, and this analgesic effect is mediated by exendin 20–29 directly inhibiting CAP-induced TRPV1 activation but not proton-induced TRPV1
activation. To determine the potential effects of exendin 20–29 on GLP-1R and glucose regulation, we performed an acute IPGTT in 8-week-old mice. As the GLP-1 analog exendin-4 (10 μg/kg)
lowers blood glucose levels47, the same amount (10 μg/kg) of exendin 20–29 was administered intraperitoneally. After 15 min of exendin 20–29 or exendin-4 administration, a glucose solution
(2 g/kg) was intraperitoneally injected (at 0 min; Fig. 8f). In contrast to exendin-4, which significantly lowered the blood glucose level from 15 to 120 min, exendin 20–29 and the control
(without pretreatment) did not change the glucose level during the experiment (Fig. 8f). This result indicates that exendin 20–29 is less likely to interact with GLP-1R (Fig. 8g). Moreover,
if the exendin 20–29 mass effective for pain relief (20 μg) was intraperitoneally injected into wild-type mice without glucose treatment, changes in blood glucose levels were not detected.
This finding indicates that exendin 20–29 does not activate GLP-1R (Supplementary Fig. 8). EXENDIN 20–29 ALLEVIATES INFLAMMATORY AND NEUROPATHIC CHRONIC PAIN BEHAVIORS Exendin 20–29 was
further assessed for its efficacy in alleviating inflammatory and neuropathic chronic pain behaviors. In the CFA-induced chronic inflammatory pain model, mice with CFA-induced inflammation
presented prolonged heat hyperalgesia following intraplantar vehicle administration. However, a single intraplantar administration of 20 or 50 μg of exendin 20–29 effectively alleviated heat
hyperalgesia, with complete recovery observed by Day 8 (Supplementary Fig. 9a). Similarly, in the von Frey test, mice injected with intraplantar vehicle after CFA induction displayed slow
recovery of mechanical allodynia. Nevertheless, a single intraplantar administration of 20 or 50 μg of exendin 20–29 alleviated mechanical allodynia (Supplementary Fig. 9b). Additionally,
the administration of exendin 20–29 resulted in a reduction in paw swelling induced by CFA (Supplementary Fig. 9c). Moreover, in the SNI-induced chronic neuropathic pain model, heat
hyperalgesia was alleviated in mice challenged with SNI and administered intraperitoneal injections of 50 μg of exendin 20–29, with effects observed as early as 1 hour postadministration and
peaking at 2 hours compared with those in the vehicle-administered group (Supplementary Fig. 9d). These findings highlight the therapeutic potential of exendin 20–29 as an analgesic agent
for chronic pain management. DISCUSSION Our study identified a novel function for GLP-1-derived peptides in providing pain relief. We specifically found that exendin 20–29 inhibits TRPV1
activity in sensory neurons in a direct and mode-specific manner, reducing pain behaviors without noticeable adverse effects. This discovery is particularly relevant in the context of the
ingestion analgesia phenomenon, which is related to the suppression of noxious heat-induced withdrawal behaviors in rats during the consumption of chocolate or sweet liquids, in contrast
with salt ingestion3,4,5,6,7. A related study revealed that the peripheral administration of D-glucose significantly enhances morphine-mediated antinociception, suggesting a synergistic
interaction between the glucose and opioid pathways48. Moreover, their findings indicated that D-glucose alone can induce significant antinociceptive responses, independent of opioid
medications. These findings suggest a broader role for glucose in pain modulation, which may inform potential therapies targeting both metabolic and pain pathways. Our primary aim was to
dissect the role of the incretin peptide hormone GLP-1 in this context by systemically administering GLP-1 and glucose via intraperitoneal injection. Our findings demonstrated that both
natural and synthetic GLP-1 administration altered heat sensitivity and that local administration of GLP-1 notably reduced heat sensitivity and CAP-induced spontaneous pain behavior. These
findings suggest that GLP-1 and its metabolites might modulate TRPV1 activity, which plays a pivotal role in heat pain perception19,49. Moreover, our study revealed that GLP-1 analogs,
including GLP-1, liraglutide, and exendin-4, along with the GLP-1 antagonist exendin 9–39, derived from exendin-4, which shares 53% sequence homology with native GLP-139, effectively
inhibited CAP-induced TRPV1 activation in mouse DRG neurons. Despite lacking the exendin 20–29 sequence RLFIEWLKNG, GLP-1 and liraglutide exhibit analgesic effects, potentially through
direct interactions with the TRPV1 receptor. These interactions may be facilitated by sequence homology with peptides such as exenatide and exendin 9–39, which incorporate segments of the
exendin sequence. Owing to the rapid degradation of GLP-1 by dipeptidyl peptidase type 4 (DPP-4), it is structurally modified into more stable GLP-1R agonists, such as liraglutide,
exenatide, and dulaglutide, which share 97%, 53%, and 90% sequence homology, respectively12. This structural modification suggests that similar analgesic effects can be mediated by these
GLP-1 analogs, even in the absence of the exendin 20–29 sequence. This concept is further illustrated in Supplementary Fig. 6, which highlights the potential binding sites for each GLP-1
analog in orange, emphasizing their role in pain modulation. The potential interaction between TRPV1 and GLP-1-derived peptides has been elucidated through structural models. Our results
indicate that these peptides modulate TRPV1 function via the extracellular membrane, effectively suppressing activation by CAP without impacting proton-related activation (Supplementary Fig.
10). These findings suggest that the peptides bind to a location that can inhibit the structural changes in TRPV1 caused by capsaicin binding but do not affect the changes caused by salt
bridge interactions between proton-sensing residues50. Although the exact binding sites remain unclear and have not been directly confirmed by structural analyses, these findings enhance our
understanding of how GLP-1-related peptides can selectively modulate TRPV1 without broad systemic effects. Unlike BCTC, exendin 9–39 also mitigated both thermal and mechanical nociceptive
behaviors in mice without inducing hyperthermia, indicating that it is a viable analgesic without the common adverse effects associated with existing pain management strategies, such as
opioids and traditional TRPV1 antagonists32,42,51,52,53. Therefore, our study indicated that GLP-1-derived peptides offer effective pain relief without these significant adverse effects,
suggesting their potential as safer alternatives for chronic pain management. Previous studies have shown that GLP-1 analogs can prevent or improve diabetic neuropathy in animal
models54,55,56,57. In addition to their neuroprotective role, GLP-1 analogs have also been studied for their influence on spinal excitatory synaptic transmission under pathophysiological
conditions such as neuropathic pain. However, previous studies have not investigated GLP-1 analogs and exendin 9–39 for pain transmission at peripheral sites, especially in small-sized
nociceptive DRG neurons, where the TRPV1 channel is crucial in chronic pain-induced mechanical allodynia and thermal hyperalgesia58,59,60,61. Recent studies have significantly enhanced our
understanding of the role of GLP-1 beyond diabetes management, revealing its substantial involvement in pain modulation. For example, research has shown that the GLP-1 receptor agonist
liraglutide can alleviate pain hypersensitivity in chronic migraine models by stimulating interleukin-10, suggesting its potential to modulate central sensitization—a crucial mechanism in
chronic pain62. Additionally, GLP-1 receptor agonists have demonstrated dual benefits in reducing headache symptoms and promoting weight loss in patients with idiopathic intracranial
hypertension, offering therapeutic advantages for conditions involving both metabolic dysregulation and chronic pain63. Furthermore, GLP-1 and glucose-dependent insulinotropic polypeptides
have been shown to induce substance P release from sensory nerves expressing TRPV1 and TRPA1, broadening the potential pain modulation mechanisms of GLP-1 and suggesting their application in
peripheral pain management64. Collectively, these findings advance the pharmacological landscape of GLP-1 and its derived peptides, suggesting that they are versatile agents for
comprehensive treatment strategies for chronic pain syndromes linked with metabolic disorders. To identify the mechanism behind the analgesic effects of GLP-1 analogs and exendin 9–39, we
utilized a cell line lacking GLP-1R to transfect the rat TRPV1 plasmid. Both GLP-1 analogs and exendin 9–39 were found to reduce CAP-induced inward currents and calcium influx through the
TRPV1 channel. Determination of the IC50 values for these compounds in CHO K1 cells expressing human TRPV1 revealed that exendin 9–39 had the lowest IC50 value for inhibiting CAP-induced
TRPV1 activation. These findings suggest that exendin 9–39 directly interacts with TRPV1, independent of GLP-1R, to inhibit its activation. This interaction was further corroborated by
pull-down assays and confocal imaging, indicating direct binding between exendin 9–39 and TRPV1 channels. Our electrophysiological analyses revealed that exendin 9–39 likely binds to the
extracellular side of TRPV1, offering selective and potent antagonism for pain relief. This finding is particularly significant, as CAP is known to bind to the transmembrane domains of TRPV1
channels60, suggesting that exendin 9–39 is a selective and potent antagonist for pain relief without competitively displacing CAP at its binding site. Although the intraperitoneal
administration of exendin 9–39 did not cause adverse hyperthermia effects, a critical aspect of developing TRPV1 antagonists is minimizing interference with TRPV1 activation by protons and
hence identifying mode-selective antagonists32. Unlike its influence on CAP-induced TRPV1 activation, exendin 9–39 did not block or potentiate proton-induced channel activity, even at a
concentration approximately 30 times greater than its IC50 value for inhibiting CAP-induced TRPV1 activity, indicating mode-specific inhibition of exendin 9–39. Critical to our analysis was
the examination of the half-maximal effective concentration values for GLP-1 analogs to activate GLP-1R, which are known to be at picomolar levels65. To exclude any potential effects of
exendin 9–39 on insulin secretion via GLP-1R interaction, we analyzed key protein sequences involved in TRPV1 inhibition. Three distinct fragments of exendin 9–39 were found to reduce
CAP-induced inward currents and calcium influx through TRPV1. Their common sequence, exendin 20–29, specifically inhibited CAP-induced TRPV1 activation without affecting proton-induced
activation and similarly alleviated CAP-induced nociceptive behaviors. To ensure that exendin 20–29 does not interact with GLP-1R, we utilized an acute IPGTT. Unlike exendin-4, exendin 20–29
did not affect blood glucose levels, supporting its lack of GLP-1R interaction. This finding was further validated by the absence of blood glucose changes in wild-type mice following the
administration of exendin 20–29 at effective doses, in contrast with the hypoglycemic effect observed with exendin-4. Although our results highlight exendin 20–29 as a promising candidate
for developing safer peptide analgesics targeting TRPV1, the translation of these findings into clinical practice requires further investigation. This includes a thorough examination of the
pharmacokinetics and dynamics in human physiology in comparison with the animal models used in our study. Given the increasing interest in peptide drugs because of their low production
costs, high potency, selectivity, and biological stability, our research underscores the potential of finding a peptide analgesic that precisely targets and modulates TRPV1 without causing
adverse effects. This represents a promising strategy in the quest for safer and more effective pain management solutions, signaling a potential shift toward innovative analgesic strategies
that prioritize both efficacy and patient safety. REFERENCES * Meleger, A. L., Froude, C. K. & Walker, J. 3rd. Nutrition and eating behavior in patients with chronic pain receiving
long-term opioid therapy. _PM R._ 6, 7–12.e11 (2014). Article PubMed Google Scholar * Amy Janke, E. & Kozak, A. T. “The more pain I have, the more I want to eat”: obesity in the
context of chronic pain. _Obesity_ 20, 2027–2034 (2012). Article CAS PubMed Google Scholar * Foo, H. & Mason, P. Sensory suppression during feeding. _Proc. Natl Acad. Sci. USA_ 102,
16865–16869 (2005). Article CAS PubMed PubMed Central Google Scholar * Casey, K. L. & Morrow, T. J. Nocifensive responses to cutaneous thermal stimuli in the cat: stimulus-response
profiles, latencies, and afferent activity. _J. Neurophysiol._ 50, 1497–1515 (1983). Article CAS PubMed Google Scholar * Wylie, L. M. & Gentle, M. J. Feeding-induced tonic pain
suppression in the chicken: reversal by naloxone. _Physiol. Behav._ 64, 27–30 (1998). Article CAS PubMed Google Scholar * Foo, H., Crabtree, K., Thrasher, A. & Mason, P. Eating is a
protected behavior even in the face of persistent pain in male rats. _Physiol. Behav._ 97, 426–429 (2009). Article CAS PubMed PubMed Central Google Scholar * Foo, H. & Mason, P.
Analgesia accompanying food consumption requires ingestion of hedonic foods. _J. Neurosci._ 29, 13053–13062 (2009). Article CAS PubMed PubMed Central Google Scholar * Ossipov, M. H.,
Dussor, G. O. & Porreca, F. Central modulation of pain. _J. Clin. Invest._ 120, 3779–3787 (2010). Article CAS PubMed PubMed Central Google Scholar * Fehmann, H. C., Goke, R. &
Goke, B. Cell and molecular biology of the incretin hormones glucagon-like peptide-I and glucose-dependent insulin releasing polypeptide. _Endocr. Rev._ 16, 390–410 (1995). Article CAS
PubMed Google Scholar * Lim, G. E. & Brubaker, P. L. Glucagon-like peptide 1 secretion by the L-cell—the view from within. _Diabetes_ 55, S70–S77 (2006). Article CAS Google Scholar
* Underwood, C. R. et al. Crystal structure of glucagon-like peptide-1 in complex with the extracellular domain of the glucagon-like peptide-1 receptor. _J. Biol. Chem._ 285, 723–730 (2010).
Article CAS PubMed Google Scholar * Brown, E., Cuthbertson, D. J. & Wilding, J. P. Newer GLP-1 receptor agonists and obesity-diabetes. _Peptides_ 100, 61–67 (2018). Article CAS
PubMed Google Scholar * Gong, N. et al. Activation of spinal glucagon-like peptide-1 receptors specifically suppresses pain hypersensitivity. _J. Neurosci._ 34, 5322–5334 (2014). Article
PubMed PubMed Central Google Scholar * Jia, Y., Gong, N., Li, T. F., Zhu, B. & Wang, Y. X. Peptidic exenatide and herbal catalpol mediate neuroprotection via the hippocampal GLP-1
receptor/beta-endorphin pathway. _Pharm. Res._ 102, 276–285 (2015). Article CAS Google Scholar * Ma, L. et al. Spinal microglial beta-endorphin signaling mediates IL-10 and
exenatide-induced inhibition of synaptic plasticity in neuropathic pain. _CNS Neurosci. Ther._ 27, 1157–1172 (2021). Article CAS PubMed PubMed Central Google Scholar * Rebosio, C.,
Balbi, M., Passalacqua, M., Ricciarelli, R. & Fedele, E. Presynaptic GLP-1 receptors enhance the depolarization-evoked release of glutamate and GABA in the mouse cortex and hippocampus.
_Biofactors_ 44, 148–157 (2018). Article CAS PubMed Google Scholar * Wu, H. Y., Mao, X. F., Fan, H. & Wang, Y. X. p38beta Mitogen- activated protein kinase signaling mediates
exenatide-stimulated microglial beta-endorphin expression. _Mol. Pharm._ 91, 451–463 (2017). Article CAS Google Scholar * Wu, H. Y., Tang, X. Q., Mao, X. F. & Wang, Y. X. Autocrine
interleukin-10 mediates glucagon-like peptide-1 receptor-induced spinal microglial beta-endorphin expression. _J. Neurosci._ 37, 11701–11714 (2017). Article CAS PubMed PubMed Central
Google Scholar * Caterina, M. J. et al. Impaired nociception and pain sensation in mice lacking the capsaicin receptor. _Science_ 288, 306–313 (2000). Article CAS PubMed Google Scholar
* Koivisto, A. P., Belvisi, M. G., Gaudet, R. & Szallasi, A. Advances in TRP channel drug discovery: from target validation to clinical studies. _Nat. Rev. Drug Discov._ 21, 41–59
(2022). Article CAS PubMed Google Scholar * Fukuoka, T. et al. VR1, but not P2X(3), increases in the spared L4 DRG in rats with L5 spinal nerve ligation. _Pain_ 99, 111–120 (2002).
Article CAS PubMed Google Scholar * Kanai, Y., Nakazato, E., Fujiuchi, A., Hara, T. & Imai, A. Involvement of an increased spinal TRPV1 sensitization through its up-regulation in
mechanical allodynia of CCI rats. _Neuropharmacology_ 49, 977–984 (2005). Article CAS PubMed Google Scholar * Yamamoto, W., Sugiura, A., Nakazato-Imasato, E. & Kita, Y.
Characterization of primary sensory neurons mediating static and dynamic allodynia in rat chronic constriction injury model. _J. Pharm. Pharm._ 60, 717–722 (2008). Article CAS Google
Scholar * Gavva, N. R. et al. The vanilloid receptor TRPV1 is tonically activated in vivo and involved in body temperature regulation. _J. Neurosci._ 27, 3366–3374 (2007). Article CAS
PubMed PubMed Central Google Scholar * Steiner, A. A. et al. Nonthermal activation of transient receptor potential vanilloid-1 channels in abdominal viscera tonically inhibits autonomic
cold-defense effectors. _J. Neurosci._ 27, 7459–7468 (2007). Article CAS PubMed PubMed Central Google Scholar * Swanson, D. M. et al. Identification and biological evaluation of
4-(3-trifluoromethylpyridin-2-yl)piperazine-1-carboxylic acid (5-trifluoromethylpyridin-2-yl)amide, a high affinity TRPV1 (VR1) vanilloid receptor antagonist. _J. Med. Chem._ 48, 1857–1872
(2005). Article CAS PubMed Google Scholar * Gavva, N. R. et al. Pharmacological blockade of the vanilloid receptor TRPV1 elicits marked hyperthermia in humans. _Pain_ 136, 202–210
(2008). Article CAS PubMed Google Scholar * Manitpisitkul, P. et al. TRPV1 antagonist JNJ-39439335 (mavatrep) demonstrates proof of pharmacology in healthy men: a first-in-human,
double-blind, placebo-controlled, randomized, sequential group study. _Pain. Rep._ 1, e576 (2016). Article PubMed PubMed Central Google Scholar * Othman, A. A., Nothaft, W., Awni, W. M.
& Dutta, S. Effects of the TRPV1 antagonist ABT-102 on body temperature in healthy volunteers: pharmacokinetic/pharmacodynamic analysis of three phase 1 trials. _Br. J. Clin. Pharm._ 75,
1029–1040 (2013). Article CAS Google Scholar * Round, P., Priestley, A. & Robinson, J. An investigation of the safety and pharmacokinetics of the novel TRPV1 antagonist XEN-D0501 in
healthy subjects. _Br. J. Clin. Pharm._ 72, 921–931 (2011). Article CAS Google Scholar * Rowbotham, M. C. et al. Oral and cutaneous thermosensory profile of selective TRPV1 inhibition by
ABT-102 in a randomized healthy volunteer trial. _Pain_ 152, 1192–1200 (2011). Article CAS PubMed Google Scholar * Garami, A. et al. TRPV1 antagonists that cause hypothermia, instead of
hyperthermia, in rodents: compounds’ pharmacological profiles, in vivo targets, thermoeffectors recruited and implications for drug development. _Acta Physiol._ 223, e13038 (2018). Article
CAS Google Scholar * Lehto, S. G. et al. Antihyperalgesic effects of (R,E)-N-(2-hydroxy-2,3-dihydro-1H-inden-4-yl)-3-(2-(piperidin-1-yl)-4-(trifluoromethyl)phenyl)-acrylamide (AMG8562), a
novel transient receptor potential vanilloid type 1 modulator that does not cause hyperthermia in rats. _J. Pharm. Exp. Ther._ 326, 218–229 (2008). Article CAS Google Scholar * Mills, C.
et al. Effects of the transient receptor potential vanilloid 1 antagonist A-425619 on body temperature and thermoregulation in the rat. _Neuroscience_ 156, 165–174 (2008). Article CAS
PubMed Google Scholar * Garami, A. et al. Contributions of different modes of TRPV1 activation to TRPV1 antagonist-induced hyperthermia. _J. Neurosci._ 30, 1435–1440 (2010). Article CAS
PubMed PubMed Central Google Scholar * Rahman, M. M., Jo, H. J., Park, C. K. & Kim, Y. H. Diosgenin exerts analgesic effects by antagonizing the selective inhibition of transient
receptor potential vanilloid 1 in a mouse model of neuropathic pain. _Int. J. Mol. Sci._ 23, 15854 (2022). Article CAS PubMed PubMed Central Google Scholar * Holst, J. J. & Gromada,
J. Role of incretin hormones in the regulation of insulin secretion in diabetic and nondiabetic humans. _Am. J. Physiol. Endocrinol. Metab._ 287, E199–E206 (2004). Article CAS PubMed
Google Scholar * Schirra, J. et al. Exendin(9-39)amide is an antagonist of glucagon-like peptide-1(7-36)amide in humans. _J. Clin. Invest._ 101, 1421–1430 (1998). Article CAS PubMed
PubMed Central Google Scholar * Deacon, C. F. Therapeutic strategies based on glucagon-like peptide 1. _Diabetes_ 53, 2181–2189 (2004). Article CAS PubMed Google Scholar * Valenzano,
K. J. et al. N-(4-tertiarybutylphenyl)-4-(3-chloropyridin-2-yl)tetrahydropyrazine -1(2H)-carbox-amide (BCTC), a novel, orally effective vanilloid receptor 1 antagonist with analgesic
properties: I. in vitro characterization and pharmacokinetic properties. _J. Pharm. Exp. Ther._ 306, 377–386 (2003). Article CAS Google Scholar * Gavva, N. R. et al. Repeated
administration of vanilloid receptor TRPV1 antagonists attenuates hyperthermia elicited by TRPV1 blockade. _J. Pharm. Exp. Ther._ 323, 128–137 (2007). Article CAS Google Scholar * Garami,
A. et al. Hyperthermia induced by transient receptor potential vanilloid-1 (TRPV1) antagonists in human clinical trials: insights from mathematical modeling and meta-analysis. _Pharm.
Ther._ 208, 107474 (2020). Article CAS Google Scholar * Luo, H., Cheng, J., Han, J. S. & Wan, Y. Change of vanilloid receptor 1 expression in dorsal root ganglion and spinal dorsal
horn during inflammatory nociception induced by complete Freund’s adjuvant in rats. _Neuroreport_ 15, 655–658 (2004). Article CAS PubMed Google Scholar * Vilceanu, D., Honore, P., Hogan,
Q. H. & Stucky, C. L. Spinal nerve ligation in mouse upregulates TRPV1 heat function in injured IB4-positive nociceptors. _J. Pain._ 11, 588–599 (2010). Article CAS PubMed Google
Scholar * Gavva, N. R. et al. Proton activation does not alter antagonist interaction with the capsaicin-binding pocket of TRPV1. _Mol. Pharm._ 68, 1524–1533 (2005). Article CAS Google
Scholar * Sedman, T., Krass, M., Runkorg, K., Vasar, E. & Volke, V. Tolerance develops toward GLP-1 receptor agonists’ glucose-lowering effect in mice. _Eur. J. Pharm._ 885, 173443
(2020). Article CAS Google Scholar * Yamamoto, R. T., Foulds-Mathes, W. & Kanarek, R. B. Antinociceptive actions of peripheral glucose administration. _Pharm. Biochem. Behav._ 117,
34–39 (2014). Article CAS Google Scholar * Davis, J. B. et al. Vanilloid receptor-1 is essential for inflammatory thermal hyperalgesia. _Nature_ 405, 183–187 (2000). Article CAS PubMed
Google Scholar * Mercadante, S., Arcuri, E. & Santoni, A. Opioid- induced tolerance and hyperalgesia. _CNS Drugs_ 33, 943–955 (2019). Article CAS PubMed Google Scholar * Wang, Z.
et al. Anti-PD-1 treatment impairs opioid antinociception in rodents and nonhuman primates. _Sci. Transl. Med._ 12, eaaw6471 (2020). Article CAS PubMed PubMed Central Google Scholar *
Szallasi, A., Cortright, D. N., Blum, C. A. & Eid, S. R. The vanilloid receptor TRPV1: 10 years from channel cloning to antagonist proof-of-concept. _Nat. Rev. Drug Discov._ 6, 357–372
(2007). Article CAS PubMed Google Scholar * Fujita, S. et al. Exenatide facilitates recovery from oxaliplatin-induced peripheral neuropathy in rats. _PLoS ONE_ 10, e0141921 (2015).
Article PubMed PubMed Central Google Scholar * Himeno, T. et al. Beneficial effects of exendin-4 on experimental polyneuropathy in diabetic mice. _Diabetes_ 60, 2397–2406 (2011). Article
CAS PubMed PubMed Central Google Scholar * Kan, M., Guo, G., Singh, B., Singh, V. & Zochodne, D. W. Glucagon-like peptide 1, insulin, sensory neurons, and diabetic neuropathy. _J.
Neuropathol. Exp. Neurol._ 71, 494–510 (2012). Article CAS PubMed Google Scholar * Liu, W. J. et al. Neuroprotective effect of the glucagon-like peptide-1 receptor agonist, synthetic
exendin-4, in streptozotocin-induced diabetic rats. _Br. J. Pharm._ 164, 1410–1420 (2011). Article CAS Google Scholar * Ji, R. R., Samad, T. A., Jin, S. X., Schmoll, R. & Woolf, C. J.
p38 MAPK activation by NGF in primary sensory neurons after inflammation increases TRPV1 levels and maintains heat hyperalgesia. _Neuron_ 36, 57–68 (2002). Article CAS PubMed Google
Scholar * Yu, L. et al. The role of TRPV1 in different subtypes of dorsal root ganglion neurons in rat chronic inflammatory nociception induced by complete Freund’s adjuvant. _Mol. Pain._
4, 61 (2008). Article PubMed PubMed Central Google Scholar * Yang, F. & Zheng, J. Understand spiciness: mechanism of TRPV1 channel activation by capsaicin. _Protein Cell_ 8, 169–177
(2017). Article CAS PubMed PubMed Central Google Scholar * Carlton, S. M. & Coggeshall, R. E. Peripheral capsaicin receptors increase in the inflamed rat hindpaw: a possible
mechanism for peripheral sensitization. _Neurosci. Lett._ 310, 53–56 (2001). Article CAS PubMed Google Scholar * Jing, F., Zou, Q. & Pu, Y. GLP-1R agonist liraglutide attenuates pain
hypersensitivity by stimulating IL-10 release in a nitroglycerin-induced chronic migraine mouse model. _Neurosci. Lett._ 812, 137397 (2023). Article CAS PubMed Google Scholar * Krajnc,
N. et al. Treatment with GLP-1 receptor agonists is associated with significant weight loss and favorable headache outcomes in idiopathic intracranial hypertension. _J. Headache Pain._ 24,
89 (2023). Article CAS PubMed PubMed Central Google Scholar * Mayer, F., Gunawan, A. L., Tso, P. & Aponte, G. W. Glucagon-like peptide 1 and glucose-dependent insulinotropic
polypeptide stimulate release of substance P from TRPV1- and TRPA1-expressing sensory nerves. _Am. J. Physiol. Gastrointest. Liver Physiol._ 319, G23–G35 (2020). Article CAS PubMed PubMed
Central Google Scholar * Mayendraraj, A., Rosenkilde, M. M. & Gasbjerg, L. S. GLP-1 and GIP receptor signaling in beta cells—a review of receptor interactions and co-stimulation.
_Peptides_ 151, 170749 (2022). Article CAS PubMed Google Scholar * Jumper, J. et al. Highly accurate protein structure prediction with AlphaFold. _Nature_ 596, 583–589 (2021). Article
CAS PubMed PubMed Central Google Scholar * Zhang, K., Julius, D. & Cheng, Y. Structural snapshots of TRPV1 reveal mechanism of polymodal functionality. _Cell_ 184, 5138–5150.e5112
(2021). Article CAS PubMed PubMed Central Google Scholar Download references ACKNOWLEDGEMENTS This study was supported by the Bio & Medical Technology Development Program of the
National Research Foundation of Korea (NRF- 2022M3E5E8081191 to C.-K.P.) and (NRF- 2021R1A5A2030333 to Y.H.K.), both of which are funded by the Korean government. We also acknowledge the
support of the U.S. National Institutes of Health (NS113243 to T.B.). AUTHOR INFORMATION Author notes * These authors contributed equally: Eun Jin Go, Sung-Min Hwang, Hyunjung Jo. AUTHORS
AND AFFILIATIONS * Gachon Pain Center and Department of Physiology, College of Medicine, Gachon University, Incheon, 21999, Republic of Korea Eun Jin Go, Sung-Min Hwang, Hyunjung Jo, Md.
Mahbubur Rahman, Jaeik Park, Yong Ho Kim & Chul-Kyu Park * Department of Anesthesiology and Pain Medicine, Gil Medical Center, Gachon University, Incheon, 21565, Republic of Korea Ji
Yeon Lee & Youn Yi Jo * Lee Gil Ya Cancer and Diabetes Institute Gachon University, Incheon, 21999, Republic of Korea Byung-Gil Lee & YunJae Jung * Pain Research Center, Department
of Anesthesiology, University of Cincinnati Medical Center, Cincinnati, OH, USA Temugin Berta Authors * Eun Jin Go View author publications You can also search for this author inPubMed
Google Scholar * Sung-Min Hwang View author publications You can also search for this author inPubMed Google Scholar * Hyunjung Jo View author publications You can also search for this
author inPubMed Google Scholar * Md. Mahbubur Rahman View author publications You can also search for this author inPubMed Google Scholar * Jaeik Park View author publications You can also
search for this author inPubMed Google Scholar * Ji Yeon Lee View author publications You can also search for this author inPubMed Google Scholar * Youn Yi Jo View author publications You
can also search for this author inPubMed Google Scholar * Byung-Gil Lee View author publications You can also search for this author inPubMed Google Scholar * YunJae Jung View author
publications You can also search for this author inPubMed Google Scholar * Temugin Berta View author publications You can also search for this author inPubMed Google Scholar * Yong Ho Kim
View author publications You can also search for this author inPubMed Google Scholar * Chul-Kyu Park View author publications You can also search for this author inPubMed Google Scholar
CONTRIBUTIONS E.J.G., S.-M.H., and H.J.J. designed the experiments. E.J.G. conducted the electrophysiological and fluorescence imaging experiments and wrote the manuscript. S.-M.H. and
H.J.J. performed the calcium imaging and molecular biology experiments. M.M.R. and J.P. performed all the behavioral tests. S.-M.H., J.Y.L., Y.Y.J., B.-G.L., Y.J., T.B., Y.H.K., and C.-K.P.
critically assisted with parts of the study and revised the manuscript accordingly. Y.H.K. and C.-K.P. supervised the study and wrote the manuscript. CORRESPONDING AUTHORS Correspondence to
Yong Ho Kim or Chul-Kyu Park. ETHICS DECLARATIONS COMPETING INTERESTS The authors declare no competing interests. ADDITIONAL INFORMATION PUBLISHER’S NOTE Springer Nature remains neutral with
regard to jurisdictional claims in published maps and institutional affiliations. SUPPLEMENTARY INFORMATION SUPPLEMENTARY INFORMATION RIGHTS AND PERMISSIONS OPEN ACCESS This article is
licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give
appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in
this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative
Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a
copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. Reprints and permissions ABOUT THIS ARTICLE CITE THIS ARTICLE Go, E.J., Hwang, SM., Jo, H. _et al._ GLP-1 and its
derived peptides mediate pain relief through direct TRPV1 inhibition without affecting thermoregulation. _Exp Mol Med_ 56, 2449–2464 (2024). https://doi.org/10.1038/s12276-024-01342-8
Download citation * Received: 07 April 2024 * Revised: 07 April 2024 * Accepted: 13 August 2024 * Published: 01 November 2024 * Issue Date: November 2024 * DOI:
https://doi.org/10.1038/s12276-024-01342-8 SHARE THIS ARTICLE Anyone you share the following link with will be able to read this content: Get shareable link Sorry, a shareable link is not
currently available for this article. Copy to clipboard Provided by the Springer Nature SharedIt content-sharing initiative