Measuring maladaptive avoidance: from animal models to clinical anxiety
- Select a language for the TTS:
- UK English Female
- UK English Male
- US English Female
- US English Male
- Australian Female
- Australian Male
- Language selected: (auto detect) - EN
Play all audios:

ABSTRACT Avoiding stimuli that predict danger is required for survival. However, avoidance can become maladaptive in individuals who overestimate threat and thus avoid safe situations as
well as dangerous ones. Excessive avoidance is a core feature of anxiety disorders, post-traumatic stress disorder (PTSD), and obsessive-compulsive disorder (OCD). This avoidance prevents
patients from confronting maladaptive threat beliefs, thereby maintaining disordered anxiety. Avoidance is associated with high levels of psychosocial impairment yet is poorly understood at
a mechanistic level. Many objective laboratory assessments of avoidance measure _adaptive_ avoidance, in which an individual learns to successfully avoid a truly noxious stimulus. However,
anxiety disorders are characterized by _maladaptive_ avoidance, for which there are fewer objective laboratory measures. We posit that maladaptive avoidance behavior depends on a combination
of three altered neurobehavioral processes: (1) threat appraisal, (2) habitual avoidance, and (3) trait avoidance tendency. This heterogeneity in underlying processes presents challenges to
the objective measurement of maladaptive avoidance behavior. Here we first review existing paradigms for measuring avoidance behavior and its underlying neural mechanisms in both human and
animal models, and identify how existing paradigms relate to these neurobehavioral processes. We then propose a new framework to improve the translational understanding of maladaptive
avoidance behavior by adapting paradigms to better differentiate underlying processes and mechanisms and applying these paradigms in clinical populations across diagnoses with the goal of
developing novel interventions to engage specific identified neurobehavioral targets. SIMILAR CONTENT BEING VIEWED BY OTHERS POST-TRAUMATIC STRESS DISORDER: CLINICAL AND TRANSLATIONAL
NEUROSCIENCE FROM CELLS TO CIRCUITS Article 29 March 2022 INTERSECT BETWEEN BRAIN MECHANISMS OF CONDITIONED THREAT, ACTIVE AVOIDANCE, AND REWARD Article Open access 26 February 2025 NEURAL
CIRCUIT BASIS OF PATHOLOGICAL ANXIETY Article 27 November 2024 INTRODUCTION Avoidance is a central and highly impairing feature of anxiety and related disorders, preventing individuals from
engaging fully in their lives [1]. Avoidance refers to any behavior that allows an individual to minimize exposure to stimuli or situations that are unpleasant, distressing, or threatening.
While avoiding stimuli that predict danger is required for survival, avoidance can become maladaptive when an individual avoids situations that are relatively safe, resulting in negative
consequences [2]. This maladaptive avoidance is a core feature of anxiety disorders and related conditions such as post-traumatic stress disorder (PTSD) and obsessive-compulsive disorder
(OCD) [3]. For example, avoidance is a diagnostic feature of social anxiety disorder and PTSD, and excessive worry in generalized anxiety disorder may allow patients to avoid distressing
visual imagery, negative emotional contrast, or feeling unsafe [4, 5]. Avoidance is a major driver that maintains anxiety pathology; it is an appealing coping strategy because it effectively
reduces anxiety in the short term, but can maintain anxiety in the long term by preventing patients from learning that their feared situations are, in fact, safe [6]. Reducing avoidance is,
therefore, an important treatment goal and a crucial part of interventions such as exposure therapy (e.g., [7]). However, while we have a thorough translational understanding of the neural
mechanisms underlying defensive reactions to acute threats such as fear and freezing, mechanisms underlying avoidance are comparatively unexplored, representing a major gap in our
understanding of anxiety pathology [8]. To advance our understanding of avoidance behavior and its neural mechanisms, it will be critical to objectively measure avoidance [9]. Current
clinical assessment of avoidance behavior largely relies on retrospective self-report (though novel approaches based on passive smartphone sensing are under development [10]). Due to the
often subtle and ingrained nature of avoidance, however, retrospective self-report may not be reliable. Furthermore, most clinical assessment of avoidance is disorder-specific (e.g., [11]),
and assessment of avoidance independent of excessive fear is lacking. There is, therefore, an unmet need for objective behavioral measurement of avoidance, outside the context of any one
disorder. Such measurement would facilitate comparison across individuals, across anxiety pathology, and across species, making it critical for mechanistic laboratory work as well as
trans-diagnostic application. In this paper, we will present three biobehavioral processes that may underlie maladaptive avoidance, describe how avoidance is most commonly measured in
laboratory settings in both animals and humans, and summarize what is already known about its neural mechanisms. We then discuss how existing paradigms could be modified to improve our
understanding of avoidance behavior and identify more precise novel targets for tailored intervention. This narrative review therefore complements and extends existing reviews of avoidance
mechanisms in rodents [12], avoidance learning in clinical anxiety [2, 13, 14], anticipation and uncertainty [15], and approach-avoidance conflict [16]. A NEUROBEHAVIORAL MODEL OF
MALADAPTIVE AVOIDANCE BEHAVIOR A key translational need is to understand _maladaptive_ avoidance, which we define as avoidance of a relatively safe stimulus often resulting in negative
consequences for the individual. Existing laboratory paradigms to study maladaptive avoidance have, therefore, examined avoidance in relative safety or in situations where negative
consequences follow from the avoidance behavior. However, even within each of these laboratory paradigms, observed avoidance behavior could be driven by different underlying processes.
Throughout this review, we will discuss three neurobehavioral processes that could drive maladaptive avoidance: (i) heightened threat appraisal, (ii) habitual avoidance, and (iii) trait
avoidance tendency, defined below. Note that while there is a technical distinction between the terms ‘fear’ and ‘anxiety’ related to acuity of threat, both acute and distant threats can
lead to avoidance behavior [17], and we therefore discuss studies of both fear and anxiety. By HEIGHTENED THREAT APPRAISAL, we mean a tendency to overestimate threat, leading to a higher
level of fear evoked by a stimulus. Heightened threat appraisal can, therefore, drive excessive avoidance behavior because of the relationship between fear and avoidance [18]. When
heightened threat appraisal is primarily responsible for maladaptive avoidance, the cause of the problem may therefore be upstream of the avoidance behavior itself, and the avoidance may
simply be a byproduct of heightened fear. For example, individuals with panic disorder may erroneously perceive an elevated heart rate from exercise as dangerous, and subsequently avoid
exercising. In this case, the mistaken perception of elevated heart rate as dangerous is the issue, and in a situation where elevated heart rate was indeed dangerous, the same avoidance
behavior would not be considered maladaptive. This neurobehavioral model is consistent with Mowrer’s classical two-factor theory, which proposes that avoidance is primarily motivated by high
levels of fear and reinforced by fear reduction [19, 20]. However, more contemporary work has demonstrated that fear and avoidance may not always be so tightly coupled (as reviewed in
[14]), and avoidance may be driven by other processes independent of high fear levels, such as habitual avoidance and/or trait avoidance tendency. HABITUAL AVOIDANCE may emerge when
avoidance is repeatedly reinforced over time. Avoidance habits are cue-based, non-goal-directed behaviors that are insensitive to outcomes [21], and have been particularly implicated in OCD
as one way to conceptualize compulsive behavior [22]. For example, a behavior such as excessive lock-checking may initially be driven by the goal of preventing burglary, then reinforced by
the feeling of brief relief from anxiety, and eventually become a habitual response to the cue of leaving home. In this case, the habitual avoidance behavior may no longer be goal directed,
and unlike avoidance driven by threat appraisal, habitual avoidance can become decoupled from the level of fear an individual feels. By TRAIT AVOIDANCE TENDENCY, we mean that there may be
stable individual differences such that even when experiencing comparable threat appraisal, some individuals may have an innate propensity to avoid to a greater extent. As with habitual
avoidance, trait avoidance tendency would therefore likely be decoupled from the level of fear (as in [23]). However, in contrast to habitual avoidance, trait avoidance tendency implies high
levels of avoidance broadly across situations, rather than specific to particular reinforced or over-learned behaviors. Although there are effective interventions that address threat
appraisals (e.g., cognitive therapy [24]) and habitual behavior (e.g., habit reversal [25]), assessment and treatment of broad trait avoidance tendencies remain relatively underdeveloped.
ADAPTIVE AVOIDANCE As a foundation to understanding maladaptive avoidance, we first review existing paradigms that assess adaptive avoidance, in which individuals avoid a truly noxious
stimulus. Adaptive avoidance behaviors fall into two broad categories: “active avoidance”, which is defined as performing a learned action to prevent harm; and “passive avoidance”, which is
defined as withholding a behavioral response to prevent harm [12]. In rodents, active avoidance has been traditionally studied using a “shuttle box” consisting of two chambers with metal bar
floors capable of delivering shocks. A neutral sensory cue (the conditioned stimulus, or CS; e.g., an auditory tone) is presented in one of the chambers and signals an impending electric
shock (the unconditioned stimulus, or US). Animals learn that “shuttling” to the other chamber in response to the CS prevents the shock from occurring. Active avoidance is thought to involve
two types of learning: (i) Pavlovian fear conditioning, in which animals learn the CS-US association (i.e., the sensory cue predicts shock); and (ii) instrumental conditioning, in which
animals learn that shuttling prevents shock [26, 27]. During early active avoidance training, Pavlovian processes dominate, as animals exhibit fearful freezing responses. Gradually
throughout training, instrumental processes dominate as animals learn that they can avoid the shock by shuttling. This paradigm is considered a measure of adaptive avoidance because animals
are avoiding the very real threat of an electric shock. Variations include “platform-mediated avoidance” [28], which adds a “safe” platform in one corner where the animals can step to avoid
the shock, and Sidman avoidance [29, 30], in which an aversive outcome occurs at a regular time interval instead of in response to a cue, resulting in repeated uncued avoidance behavior. A
related type of task, called aversive Pavlovian Instrumental Transfer (PIT), explicitly separates the Pavlovian and instrumental learning processes enabling investigations into the
relationship between each of these learning processes [31, 32]. The active avoidance paradigm has also been adapted for use in human research with three major differences. First, human
analogues typically do not involve moving to a different location to avoid an aversive outcome but often rely on behaviors such as a button press in computer-based tasks (though see also
[33]). Second, in human analogues the experimenter often verbally instructs the participant on the CS-US association, resulting in a minimal contribution of Pavlovian learning. When
Pavlovian learning is included in the paradigm, it usually is conducted without any opportunity to engage in avoidance behavior [34]. This has the benefit of isolating the fear and avoidance
learning components, with the drawback of fewer parallels to rodent paradigms. Third, while in rodent paradigms avoidance behavior is always learned through trial-and-error, human avoidance
paradigms can either be learned (e.g., “one of these buttons will prevent shock”; [35]) or fully instructed (e.g., “press here to prevent shock”; [36]). Rodent and human studies employing
these active avoidance tasks have converged on three critical neural structures: the medial prefrontal cortex (mPFC), the amygdala, and the striatum [28, 30, 35, 37,38,39,40]. The mPFC is
implicated in top-down control of emotion and decision-making and is thought to modulate subcortical structures such as the central and basolateral amygdala [29, 41, 42] and the ventral and
dorsal striatum [12, 43,44,45] that are implicated in valence processing and defensive behaviors including active avoidance [39, 46, 47]. Indeed, greater synchrony between mPFC and both the
amygdala and striatum predicts effective avoidance learning [30]. Amygdala-striatal circuitry is also thought to be important for avoidance learning [35], although some studies indicate that
the amygdala may no longer be required for avoidance expression after extensive training, suggesting that frontostriatal mechanisms of habitual avoidance may dominate after avoidance is
well learned [29]. However, the majority of work on frontostriatal circuitry in instrumental learning has used _positive reinforcement_ [48,49,50,51], whereas active avoidance learning can
be a form of _negative reinforcement_. Whether the frontostriatal circuit mechanisms underlying positive reinforcement are also relevant to avoidance remains an open question of critical
importance to understanding avoidance mechanisms [52]. In contrast to active avoidance, passive avoidance involves withholding a behavioral response in order to prevent harm. Passive
avoidance has traditionally been conceptualized as a readout of fear memory [53,54,55,56], and work in rodents and non-human primates have implicated the amygdala and extended amygdala in
passive defensive responding [57,58,59]. However, little is known mechanistically about passive avoidance behavior at the circuit level, likely because it is difficult to identify neural
correlates in the absence of a behavior. In addition, few studies have directly compared active and passive avoidance behavior to identify similarities and differences in neural mechanisms
(though see [45, 60] for paradigms that facilitate this comparison). MALADAPTIVE AVOIDANCE While much is known about the neural circuitry underlying _adaptive_ avoidance (i.e., avoidance of
a truly noxious stimulus), anxiety disorders, OCD, and PTSD are typically characterized by _maladaptive_ avoidance. We define maladaptive avoidance as avoidance of a relatively safe stimulus
and/or resulting in negative consequences for the individual, such as loss of reward. To model avoidance of a relatively safe stimulus, paradigms typically examine either
extinction-resistant avoidance, or generalization of avoidance to a cue perceptually similar to a conditioned stimulus. To model negative consequences from avoidance, paradigms often
introduce competing rewards that the individual must forgo to perform the avoidance behavior. Common and promising examples of paradigms that model maladaptive avoidance are reviewed below
and summarized in Table 1. EXTINCTION-RESISTANT AVOIDANCE Extinction-resistant avoidance is defined as avoidance behavior that persists even after Pavlovian extinction training in which the
CS (cue) is repeatedly presented in the absence of the US (aversive stimulus), and is maladaptive because the CS no longer predicts threat. Extinction-resistant avoidance has clear clinical
relevance because continued avoidance following exposure therapy (which is thought to work through extinction) may put patients at higher risk for relapse [61, 62]. However,
extinction-resistant avoidance could be due to more than one underlying neurobehavioral process. Extinction-resistant avoidance may be due to heightened threat appraisal, particularly if
both fear _and_ avoidance persist following extinction training. Such incomplete extinction of both fear and avoidance is more common when avoidance behavior is not prevented during
extinction training. In this case, animals continue to avoid during extinction training, thereby receiving fewer opportunities to extinguish the CS-US association and resulting in persistent
fear [63]. Alternatively, despite the often strong coupling between fear and avoidance, extinction-resistant avoidance behavior can occur even when extinction training has successfully
reduced conditioned fear responding [63, 64]. In these cases, avoidance may have become a habit and thus governed by different neural circuitry. Animal studies of extinction-resistant
avoidance behavior have largely built on active avoidance paradigms. After avoidance learning, animals undergo extinction training in which the CS is presented in the absence of the US.
Although most animals learn that there is no longer a need to avoid the CS, there are large individual differences in avoidance following extinction training, with a substantial fraction of
animals continuing to persistently avoid [63, 64]. However, it is hard to distinguish whether this extinction-resistant avoidance is simply a product of insufficient fear extinction without
measuring both fear and avoidance. Human studies of extinction-resistant avoidance have more consistently measured both fear and avoidance, allowing stronger conclusions to be drawn about
the neurobehavioral process underlying the observed avoidance behavior. For example, greater avoidance behavior has been observed in OCD patients than in healthy controls, in the absence of
fear-related differences indexed by skin conductance [65]. Furthermore, greater avoidance in OCD was only seen after a large number of avoidance trials, suggesting a habitual component to
the avoidance. In contrast, extinction-resistant avoidance behavior has been observed in the context of incomplete extinction of fear, which suggests that this persistent avoidance behavior
may be driven by continued threat appraisal [36]. Further supporting the notion that a threat-related process drives avoidance was the relationship between trait anxiety and
extinction-resistance avoidance, as well as the relatively small number of avoidance training trials in this study [36]. Another approach to disentangling the relative contribution of threat
appraisal and habit in extinction-resistant avoidance is using response prevention (i.e., removing the opportunity to avoid during extinction training) to ensure that fear responses are
completely extinguished. Rodent studies have directly compared neural correlates of persistent avoidance following extinction training with and without response prevention.
Extinction-resistant avoidance was correlated with increased activity in the prelimbic cortex and ventral striatum regardless of response prevention, but when response prevention was absent,
extinction-resistant avoidance was also correlated with impaired recruitment of regions associated with fear extinction (i.e., increased basal amygdala and decreased infralimbic cortex
activity) [63]. These results suggest that without response prevention, animals may not have sufficient opportunity to extinguish fear, and this heightened threat appraisal may be driving
persistent avoidance. In contrast, persistent avoidance following extinction _with_ response prevention may be due to habitual avoidance (though spontaneous recovery of fear or lack of cost
for continuing to perform the avoidance response may also contribute). Habitual avoidance has been particularly implicated in OCD, and clinical treatment of OCD typically involves exposure
to feared stimuli (e.g., dirty objects) combined with response prevention of compulsive avoidance behavior (e.g., hand-washing). A recent study showed that overtraining on avoidance behavior
biased rats toward habitual avoidance following extinction training with response prevention, which was associated with increased activity in frontostriatal regions [66]. This paradigm is a
promising model of habitual avoidance that may be present in OCD, and presents an opportunity for further investigation into circuit mechanisms underlying extinction-resistant avoidance.
Furthermore, at least one inactivation study in rodents suggests that extinction-resistant avoidance following response prevention depends on the lateral orbitofrontal cortex [67], which has
been implicated in both habit [68] and OCD [69]. Functional neuroimaging of adults with OCD has also implicated the caudate nucleus in persistent habitual avoidance in this population [70].
AVOIDANCE GENERALIZATION Generalization is a phenomenon in which individuals apply learning from past experiences to similar related situations or stimuli [71]. Generalization can be
adaptive by allowing us to navigate complex risks based on limited experience. However, over-generalization of avoidance behavior is maladaptive, as it leads to avoidance of related but safe
situations. Avoidance over-generalization is a core feature of anxiety and related disorders [13]. For example, someone with PTSD as a result of a car accident may not only avoid the
intersection where the crash occurred, but may generalize to avoiding driving altogether. Avoidance generalization could be driven by the underlying processes of heightened threat appraisal,
if the avoidance generalization is proportional to the generalization of fear, or by trait avoidance tendency. Although few rodent studies have examined _avoidance_ generalization, many
have focused on _fear_ generalization. These studies have shown that tonic activity of central amygdala neurons correlates with the degree of fear generalization [72], and increased
synchrony between basolateral amygdala and medial prefrontal neurons occurs in animals demonstrating less fear generalization [73]. However, little is known about whether these same circuits
regulate generalization of avoidance responses. In contrast to the limited rodent models of avoidance generalization, there has been a surge of interest in human avoidance generalization
paradigms (e.g., [74,75,76]). For example, in the virtual farmer game [77], participants first undergo Pavlovian conditioning to associate a shape (CS) with a shock. Next, participants must
move a farmer icon between images of a shed and crops, aiming to arrive at the crops before virtual birds interfere. They are given the option of a short path, which results in shock when
the CS is present, or a long path, which interferes with the goal of the game but is always safe from shock. Avoidance generalization is measured by the extent to which participants select
the long path when generalization stimuli (with some perceptual similarity to the CS but never paired with shock) are present. As with extinction-resistant avoidance, experimentally
measuring both fear and avoidance in the same tasks will help distinguish the underlying neurobehavioral processes driving avoidance generalization. In the virtual farmer paradigm,
individuals who exhibited greater fear generalization (measured by skin conductance) demonstrated greater avoidance generalization (measured by selecting the long path), suggesting that
heightened threat appraisal may be a driver of avoidance generalization in this paradigm [77]. This paradigm could also be used to identify individual differences in trait avoidance
tendency, for example, if individuals avoid a generalization stimulus in the absence of significant fear responding. Greater avoidance generalization in the virtual farmer game was
associated with individual differences in avoidant coping styles such as distraction or suppression [77], consistent with the notion that trait avoidance tendency may play a role in
avoidance generalization for some individuals. Neural mechanisms of avoidance generalization are not well understood relative to other paradigms (Table 1). Extending animal models of fear
generalization to avoidance behavior would help delineate neural mechanisms, and is an important future direction. COMPETING REWARDS Maladaptive avoidance behavior often results in negative
consequences such as loss of opportunity for positive outcomes. Thus, a common strategy to study maladaptive avoidance behavior is to pit avoidance against a competing reward, such that
avoidance results in both a decreased risk of harm _and_ a decreased potential for reward. This approach parallels clinical situations such as in social anxiety, where avoidance of social
interaction results in loss of both potential negative outcomes (e.g., being rejected) and potential positive outcomes (e.g., making friends). Avoidance in the face of competing reward could
be driven by either acquired habitual avoidance, or an innate imbalance between trait avoidance tendency and reward sensitivity. If behavior is habitual, it is by definition not
goal-directed and will persist regardless of the reward accrued. In addition, individuals may differ in the extent to which they are either predisposed to avoid and/or sensitive to the
potential for reward, either of which would result in greater avoidance decisions when competing rewards are present [78]. One example of how competing rewards can be implemented in animal
models is the platform-mediated avoidance test [28], in which animals can press a lever for a food reward as long as they are on the shock floor. When they step onto the platform to avoid
the shock, however, they are no longer able to reach the lever and therefore forgo this reward. This task has been further modified to maximize competition between avoidance and reward by
presenting the lever for reward only during presentation of the tone that signals impending shock. There were individual differences in avoidance tendency such that rats that preferred
avoidance had increased amygdala activity compared to rats that preferred reward approach, which had decreased prefrontal activity [79]. The “risk-reward interaction” task [60] also adds an
element of reward into a traditional shuttle box style apparatus, such that each chamber contains a shock floor and a reward port. This task has been used to measure neural signals related
to active avoidance and reward-seeking behaviors on separate trials but could also be adapted to make reward only accessible by crossing a shock floor. This would allow researchers to
measure how much an animal is willing to forgo reward in order to avoid, and examine if different neural mechanisms are involved when avoidance is paired with competing rewards. Variations
of the risk-reward interaction task could also be used to assess how decisions to forgo rewards relate to the acquisition of avoidance habits, and whether different neural mechanisms are
involved in this decision once avoidance has become habitual. In addition, although these paradigms have thus far been used with the fixed intensity of threats and rewards, varying the
intensity could better probe individual thresholds for maladaptive avoidance behavior. A complementary approach to these acute assays is the semi-naturalistic “closed economy” paradigm in
which animals live for several weeks in the apparatus and must forage for food in a location that also delivers random electric shocks, allowing for ethologically relevant probing of
avoidance behaviors as a reward and/or shock schedules are varied [80,81,82]. Human studies have begun investigating the impact of varying threat and reward intensities using computer games
that provide participants the opportunity to earn rewards (e.g., money) by facing negative consequences, or avoid these consequences without receiving any reward [78, 83]. While related work
has investigated _loss_ of money as a negative consequence [84, 85], we focus here on paradigms involving the _addition_ of an aversive outcome, such as viewing threatening images or
receiving the shock. In particular, recent work examined avoidance decision-making under varying combinations of shock probability and reward magnitude [83]. High anxiety individuals made
more avoidance decisions compared to low anxiety individuals as reward magnitude increased, suggesting that avoidance decisions in anxious individuals may be driven by low reward
sensitivity. Although the neural mechanisms underlying this paradigm have not been examined, functional neuroimaging studies of similar paradigms have implicated the anterior insula and
medial prefrontal cortex [16, 86]. In non-human primates, amygdala lesions increase avoidance in the presence of competing rewards [87], while stimulation of the anterior insula and ventral
striatum increases this avoidance behavior [88]. A related type of paradigm is an approach-avoidance conflict task, which presents an animal with conflicting appetitive and aversive cues
that must be simultaneously weighed. In contrast to the examples above, which use _learned_ avoidance behavior, approach-avoidance conflict tasks have traditionally examined _innate_
avoidance behavior. The most common approach-avoidance conflict assay is the elevated plus maze, which is a platform consisting of two “open” exposed arms and two “closed” arms with tall
walls that animals freely explore [89, 90]. This task capitalizes on rodents’ conflicting innate aversion to open brightly lit spaces (for risk of predation) and innate drive to explore
novel environments. There are many related tasks to study conflicting costs and benefits. For example, the T-maze measures how willing an animal is to avoid an innately aversive stimulus,
such as bright light, at the cost of losing a reward, such as food [91]. Semi-natural foraging environments that employ food rewards and robotic predators have likewise been used to study
approach-avoidance conflict behaviors [58]. In addition, human and non-human primate studies have used a joystick to capitalize on the instinct to pull desired stimuli closer and push
aversive stimuli away [92]. Due to the innate nature of the avoidance behavior in approach-avoidance conflict tasks, greater maladaptive avoidance in these tasks can be considered a measure
of trait avoidance tendency. Approach-avoidance conflict tasks require the mPFC: ventromedial aspects that project to the basolateral amygdala control open arm exploration in the elevated
plus maze [47], whereas dorsomedial aspects that project to the dorsal striatum control cost-benefit decision-making in the T-maze task as well as innate avoidance in the elevated plus maze
[91, 93]. This suggests an important role of frontostriatal projections in trait avoidance tendency. There is also some encouraging evidence that these tasks have relevance for human
anxiety. Human analogues of approach-avoidance conflict tasks have used movement tracking to measure innate avoidance versus exploration behavior in real or virtual settings [94,95,96,97].
These studies have generally found greater innate avoidance behavior in those with higher anxiety, and decreased avoidance following anxiolytic medication, validating the potential utility
of this approach for understanding anxiety disorders and trait-like individual differences. GAPS IN KNOWLEDGE Moving forward, it will be important to address four key areas: differentiating
neurobehavioral alterations, translating insights across species, testing paradigms in clinical populations, and improving diagnosis and treatment (Table 2). DIFFERENTIATING NEUROBEHAVIORAL
ALTERATIONS A major gap in the field is the lack of translational paradigms that can differentiate which neurobehavioral alterations underlie a particular individual’s maladaptive avoidance
behavior. An important next step will therefore be to develop tasks that better differentiate the processes and circuitry underlying maladaptive avoidance. Existing paradigms could be
modified to determine the extent to which fear and avoidance are correlated by including quantifiable proxies for fear (e.g., autonomic measures such as heart rate or skin conductance) and
investigating how closely they track with avoidance behavior over time. A close coupling between fear and avoidance would suggest a greater contribution of threat appraisal to the avoidance
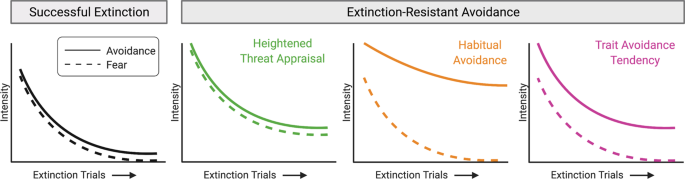
behavior, whereas lower coupling would imply that habitual avoidance and/or avoidance tendency are involved (Fig. 1). In addition, examining the temporal dynamics of avoidance could help
differentiate underlying processes. For example, because habits are less sensitive to action-outcome relationships, avoidance behavior driven by habit would be less likely to diminish during
extinction training (Fig. 1). In animal studies, differentiating between these specific underlying processes will require longitudinal tracking of both behavior and neural activity at a
resolution that has not yet been brought to the field. Greater specificity in differentiating underlying processes will also require measuring defensive behaviors beyond avoidance (e.g.,
freezing), and recording neural activity not only during avoidance expression but also during avoidance learning and extinction. Combining these behavioral measurements with
cell-type-specific and neural projection-specific circuit dissection techniques, such as optogenetics and in vivo calcium imaging, will be critical for elucidating the real-time circuit
dynamics correlated with each of these behaviors. TRANSLATING INSIGHTS ACROSS SPECIES Animal models can provide greater spatial and temporal precision in identifying neural mechanisms, while
human research can assess internal experiences (e.g., feeling anxious), and provide verbal instruction on available avoidance behaviors. Such instructed avoidance paradigms may allow for
better separation of trait avoidance tendency and habit because instructed avoidance behavior removes the possibility that repeated practice of avoidance has led to habitual responding.
However, while these unique insights are important, it is also important to align paradigms across human and animal models to better link the unique insights from each species. This should
include alignment on the precise use of terminology, such as “relief” (a clinically-relevant subjective experience) versus “shock omission” (an objectively defined event). Human and animal
paradigms will have better alignment when they involve common behavioral outputs and physiological measures during avoidance tasks. Additionally, quickly-developing technology such as
virtual reality platforms [98] and smartphone-based passive sampling [99] offer a unique ability to align tasks across species by re-creating for humans the freely moving laboratory tasks
from rodent work. Computational models that can generate hypotheses independent of the species being studied are another important approach to facilitate translation [100]. Finally, while
the majority of research has focused on rodent and human models of avoidance behavior, more work is needed to translate across non-human primate and human studies, given the advantage of
greater functional homology of the brain relative to rodent models [101, 102]. TESTING PARADIGMS IN CLINICAL POPULATIONS Another crucial gap is understanding how avoidance behavior as
defined in laboratory paradigms applies in clinical populations. It remains unclear whether the mechanisms uncovered from studies of adaptive avoidance are the same as those that go awry in
clinical conditions that are defined by maladaptive avoidance. The majority of paradigms described above have not been tested in individuals with clinical levels of anxiety and avoidance;
doing so would allow for a more nuanced characterization of maladaptive avoidance behavior within and across disorders. For example, the distinction between active and passive avoidance
strategies is commonly made in animal models, yet rarely made in describing clinical phenomena, highlighting a missing translational link. Moreover, it is not known whether mechanisms
underlying external avoidance (e.g., of electric shock) are the same as those underlying experiential avoidance (e.g., of unpleasant thoughts or emotions) common in anxiety and related
disorders. Despite large interest in trans-diagnostic features of psychopathology, it is still unclear whether avoidance involves the same neural mechanisms across disorders. For example,
does avoidance of social situations in social anxiety disorder involve the same mechanisms as avoidance of trauma reminders in PTSD? Understanding how specific neurobehavioral processes map
onto existing DSM diagnoses or trans-diagnostic features (such as worry or intolerance of uncertainty) will be an important step to bridge current clinical practice and a neuroscience-based
understanding of maladaptive avoidance behavior. Finally, it will be important for the field to develop paradigms that mimic the probabilistic nature of safety in the real world, in addition
to existing experimental laboratory paradigms that can more objectively define safety. IMPROVING DIAGNOSIS AND TREATMENT Assuming that distinct neurobehavioral processes reflect different
root causes of maladaptive avoidance behavior, an open question is whether they could someday provide a more precise and useful classification system than a current clinical practice using
DSM symptom-based classification. This goal would require reliable and valid measurement of avoidance behavior at the level of an individual, with known population norms and/or clinical cut
points. While some studies have begun to examine construct validity [36, 53], psychometric properties of avoidance paradigms have largely not been established (for a notable exception, see
[103]). Psychometrically sound tasks will be essential for translation from research to the clinical application [104], with test-retest reliability of particular importance for paradigms
assessing constructs that should be stable over time, such as trait avoidance tendency. Using a classification system based on reliable and valid measurement of underlying neurobehavioral
alterations, we may be able to better tailor interventions to target specific avoidance problems unique to each patient. Although the current gold-standard treatment approach for anxiety and
related disorders is exposure therapy, this treatment is not successful for many patients [105]. Exposure therapy is thought to work through fear extinction [106]; this extinction-based
view of exposure therapy suggests that exposure therapy will be most effective in addressing avoidance behaviors driven by heightened threat appraisal. However, it is unclear whether
exposure therapy can also effectively modify avoidance behavior that has become decoupled from fear, as with trait avoidance tendency or habitual avoidance, or whether maladaptive avoidance
behavior driven by these processes could be better addressed by operant reinforcement principles rather than Pavlovian and inhibitory learning processes. For example, one hypothesis is that
interventions such as acceptance and commitment therapy (ACT) that focus on approach towards values-based behaviors [62, 107] may be more effective treatments for trait avoidance. While one
study has found that avoidance is associated with more success in CBT than ACT [9], the neurobehavioral process underlying this avoidance is unclear. Ultimately, just as insights from the
laboratory paradigm of fear extinction have improved exposure therapy [106], laboratory paradigms that can measure and manipulate maladaptive avoidance have the potential to identify and
optimize new treatment approaches to more directly target additional root causes of maladaptive avoidance behavior. CONCLUSION Avoidance is a central and trans-diagnostic mechanism that
leads to impairment across anxiety pathology, including OCD and PTSD. Reducing maladaptive avoidance is an important treatment goal and a necessary part of evidence-based interventions such
as exposure therapy. We propose that maladaptive avoidance may be caused by alterations in three distinct neurobehavioral processes: (i) threat appraisal, (ii) habitual avoidance, and (iii)
trait avoidance tendency. Existing paradigms to measure maladaptive avoidance have examined extinction-resistant avoidance, avoidance generalization, and avoidance in the face of competing
rewards. An important next step for the field will be to adapt these paradigms to more precisely differentiate which neurobehavioral alterations underlie a particular individual’s
maladaptive avoidance behavior and use these paradigms to understand how specific neurobehavioral processes map onto clinical diagnoses. Ultimately, this work has the potential to lead to
new interventions to more effectively treat maladaptive avoidance behavior in anxiety and related disorders. REFERENCES * Salters-Pedneault K, Tull MT, Roemer L. The role of avoidance of
emotional material in the anxiety disorders. Appl Prev Psychol. 2004;11:95–114. Article Google Scholar * Arnaudova I, Kindt M, Fanselow M, Beckers T. Pathways towards the proliferation of
avoidance in anxiety and implications for treatment. Behav Res Ther. 2017;96:3–13. Article PubMed Google Scholar * Hofmann SG, Hay AC. Rethinking avoidance: toward a balanced approach to
avoidance in treating anxiety disorders. J Anxiety Disord. 2018;55:14–21. Article PubMed PubMed Central Google Scholar * Sibrava NJ, Borkovec TD. The Cognitive Avoidance Theory of Worry.
In: Davey GCL, Wells A. editors. Worry and its psychological disorders: Theory, assessment and treatment. Wiley Publishing; 2006. p. 239–56. * Newman MG, Llera SJ. A novel theory of
experiential avoidance in generalized anxiety disorder: a review and synthesis of research supporting a contrast avoidance model of worry. Clin Psychol Rev. 2011;31:371–82. Article PubMed
PubMed Central Google Scholar * Thwaites R, Freeston MH. Safety-seeking behaviours: fact or function? How can we clinically differentiate between safety behaviours and adaptive coping
strategies across anxiety disorders? Behav Cogn Psychother. 2005;33:177–88. Article Google Scholar * Hedman-Lagerlöf M, Andersson E, Hedman-Lagerlöf E, Wicksell RK, Flink I, Ljótsson B.
Approach as a key for success: reduced avoidance behaviour mediates the effect of exposure therapy for fibromyalgia. Behav Res Ther. 2019;122:103478. Article PubMed Google Scholar *
Tovote P, Fadok JP, Lüthi A. Neuronal circuits for fear and anxiety. Nat Rev Neurosci. 2015;16:317–31. Article CAS PubMed Google Scholar * Mesri B, Niles AN, Pittig A, LeBeau RT, Haik E,
Craske MG. Public speaking avoidance as a treatment moderator for social anxiety disorder. J Behav Ther Exp Psychiatry. 2017;55:66–72. Article PubMed Google Scholar * Boukhechba M,
Baglione AN, Barnes LE. Leveraging mobile sensing and machine learning for personalized mental health care. Erg Des. 2020;28:18–23. Google Scholar * Heimberg RG, Horner KJ, Juster HR,
Safren SA, Brown EJ, Schneier FR, et al. Psychometric properties of the Liebowitz Social Anxiety Scale. Psychol Med. 1999;29:199–212. Article CAS PubMed Google Scholar * LeDoux JE,
Moscarello J, Sears R, Campese V. The birth, death and resurrection of avoidance: a reconceptualization of a troubled paradigm. Mol Psychiatry. 2017;22:24–36. Article CAS PubMed Google
Scholar * Pittig A, Treanor M, LeBeau RT, Craske MG. The role of associative fear and avoidance learning in anxiety disorders_ Gaps and directions for future research. Neurosci Biobehav
Rev. 2018;88:117–40. Article PubMed Google Scholar * Krypotos A-M, Effting M, Kindt M, Beckers T. Avoidance learning: a review of theoretical models and recent developments. Front Behav
Neurosci. 2015;9:189. Article PubMed PubMed Central Google Scholar * Grupe DW, Nitschke JB. Uncertainty and anticipation in anxiety: an integrated neurobiological and psychological
perspective. Nat Rev Neurosci. 2013;14:488–501. Article CAS PubMed PubMed Central Google Scholar * Kirlic N, Young J, Aupperle RL. Animal to human translational paradigms relevant for
approach avoidance conflict decision making. Behav Res Ther. 2017;96:14–29. Article PubMed PubMed Central Google Scholar * Perusini JN, Fanselow MS. Neurobehavioral perspectives on the
distinction between fear and anxiety. Learn Mem. 2015;22:417–25. Article CAS PubMed PubMed Central Google Scholar * Grant DM, Wingate LR, Rasmussen KA, Davidson CL, Slish ML,
Rhoades-Kerswill S, et al. An examination of the reciprocal relationship between avoidance coping and symptoms of anxiety and depression. J Soc Clin Psychol. 2013;32:878–96. Article Google
Scholar * Mowrer OH. Learning theory and behavior. In Hoboken NJ (editor), John Wiley & Sons; 1960. * Mowrer OH. Two-factor learning theory: summary and comment. Psychol Rev.
1951;58:350–4. Article CAS PubMed Google Scholar * Dickinson A. Actions and habits: the development of behavioural autonomy. Philos Trans R Soc Lond B Biol Sci. 1985;308:67–78. Article
Google Scholar * Chase HW, Graur S, Versace A, Greenberg T, Bonar L, Hudak R, et al. Neural mechanisms of persistent avoidance in OCD: A novel avoidance devaluation study. Neuroimage Clin.
2020;28:102404. Article PubMed PubMed Central Google Scholar * Effting M, Salemink E, Verschuere B, Beckers T. Implicit and explicit measures of spider fear and avoidance behavior:
Examination of the moderating role of working memory capacity. J Behav Ther Exp Psychiatry. 2016;50:269–76. Article PubMed Google Scholar * Clark DA, Beck AT. Cognitive theory and therapy
of anxiety and depression: convergence with neurobiological findings. Trends Cogn Sci. 2010;14:418–24. Article PubMed Google Scholar * Bate KS, Malouff JM, Thorsteinsson ET, Bhullar N.
The efficacy of habit reversal therapy for tics, habit disorders, and stuttering: a meta-analytic review. Clin Psychol Rev. 2011;31:865–71. Article PubMed Google Scholar * Mowrer O.
Learning theory and behavior. John Wiley & Sons Inc.; 1960. * Rachman S. The passing of the two-stage theory of fear and avoidance: fresh possibilities. Behav Res Ther. 1976;14:125–31.
Article CAS PubMed Google Scholar * Bravo-Rivera C, Roman-Ortiz C, Brignoni-Perez E, Sotres-Bayon F, Quirk GJ. Neural structures mediating expression and extinction of platform-mediated
avoidance. J Neurosci. 2014;34:9736–42. Article CAS PubMed PubMed Central Google Scholar * Lazaro-Munoz G, LeDoux JE, Cain CK. Sidman instrumental avoidance initially depends on lateral
and basal amygdala and is constrained by central amygdala-mediated Pavlovian processes. Biol Psychiatry. 2010;67:1120–7. Article PubMed PubMed Central Google Scholar * Collins KA,
Mendelsohn A, Cain CK, Schiller D. Taking action in the face of threat: neural synchronization predicts adaptive coping. J Neurosci. 2014;34:14733–8. Article CAS PubMed PubMed Central
Google Scholar * Campese VD, Kim IT, Kurpas B, Branigan L, Draus C, LeDoux JE. Motivational factors underlying aversive Pavlovian-instrumental transfer. Learn Mem. 2020;27:477–82. Article
PubMed PubMed Central Google Scholar * Lewis AH, Niznikiewicz MA, Delamater AR, Delgado MR. Avoidance-based human Pavlovian-to-instrumental transfer. Eur J Neurosci. 2013;38:3740–8.
Article PubMed Google Scholar * Mobbs D, Marchant JL, Hassabis D, Seymour B, Tan G, Gray M, et al. From threat to fear: the neural organization of defensive fear systems in humans. J
Neurosci. 2009;29:12236–43. Article CAS PubMed PubMed Central Google Scholar * Lovibond PF, Saunders JC, Weidemann G, Mitchell CJ. Evidence for expectancy as a mediator of avoidance and
anxiety in a laboratory model of human avoidance learning. Q J Exp Psychol. 2008;61:1199–216. Article Google Scholar * Delgado MR. Avoiding negative outcomes: tracking the mechanisms of
avoidance learning in humans during fear conditioning. Front Behav Neurosci. 2009;3:1–9. Article Google Scholar * Vervliet B, Indekeu E. Low-cost avoidance behaviors are resistant to fear
extinction in humans. Front Behav Neurosci. 2015;9:351. Article PubMed PubMed Central Google Scholar * Diehl MM, Bravo-Rivera C, Rodriguez-Romaguera J, Pagan-Rivera PA, Burgos-Robles A,
Roman-Ortiz C, et al. Active avoidance requires inhibitory signaling in the rodent prelimbic prefrontal cortex. Elife. 2018;7:e34657. * Ramirez F, Moscarello JM, LeDoux JE, Sears RM. Active
avoidance requires a serial basal amygdala to nucleus accumbens shell circuit. J Neurosci. 2015;35:3470–7. Article CAS PubMed PubMed Central Google Scholar * Diehl MM, Iravedra-Garcia
JM, Morán-Sierra J, Rojas-Bowe G, Gonzalez-Diaz FN, Valentín-Valentín VP, et al. Divergent projections of the prelimbic cortex bidirectionally regulate active avoidance. Elife.
2020;9:e59281. * Hur J, Stockbridge MD, Fox AS, Shackman AJ. Dispositional negativity, cognition, and anxiety disorders: An integrative translational neuroscience framework. Prog Brain Res.
2019;247:375–436. Article PubMed PubMed Central Google Scholar * Jiao X, Beck KD, Myers CE, Servatius RJ, Pang KC. Altered activity of the medial prefrontal cortex and amygdala during
acquisition and extinction of an active avoidance task. Front Behav Neurosci. 2015;9:249. Article PubMed PubMed Central Google Scholar * Yu K, Garcia da Silva P, Albeanu DF, Li B.
Central amygdala somatostatin neurons gate passive and active defensive behaviors. J Neurosci. 2016;36:6488–96. Article CAS PubMed PubMed Central Google Scholar * Xiao X, Deng H, Furlan
A, Yang T, Zhang X, Hwang G-R, et al. A genetically defined compartmentalized striatal direct pathway for negative reinforcement. Cell. 2020;183:211–e20. Article CAS PubMed PubMed
Central Google Scholar * González-Salinas S, Medina AC, Alvarado-Ortiz E, Antaramian A, Quirarte GL, Prado-Alcalá RA. Retrieval of inhibitory avoidance memory induces differential
transcription of arc in striatum, hippocampus, and amygdala. Neuroscience. 2018;382:48–58. Article PubMed Google Scholar * Levita L, Hoskin R, Champi S. Avoidance of harm and anxiety: A
role for the nucleus accumbens. Neuroimage. 2012;62:189–98. Article PubMed Google Scholar * Giustino TF, Maren S. The role of the medial prefrontal cortex in the conditioning and
extinction of fear. Front Behav Neurosci. 2015;9:298. Article PubMed PubMed Central Google Scholar * Adhikari A, Lerner TN, Finkelstein J, Pak S, Jennings JH, Davidson TJ, et al.
Basomedial amygdala mediates top-down control of anxiety and fear. Nature. 2015;527:179–85. Article CAS PubMed PubMed Central Google Scholar * Ostlund SB, Balleine BW. Lesions of medial
prefrontal cortex disrupt the acquisition but not the expression of goal-directed learning. J Neurosci. 2005;25:7763–70. Article CAS PubMed PubMed Central Google Scholar * Yin HH,
Ostlund SB, Knowlton BJ, Balleine BW. The role of the dorsomedial striatum in instrumental conditioning. Eur J Neurosci. 2005;22:513–23. Article PubMed Google Scholar * Simon NW, Wood J,
Moghaddam B. Action-outcome relationships are represented differently by medial prefrontal and orbitofrontal cortex neurons during action execution. J Neurophysiol. 2015;114:3374–85. Article
CAS PubMed PubMed Central Google Scholar * Kravitz AV, Tye LD, Kreitzer AC. Distinct roles for direct and indirect pathway striatal neurons in reinforcement. Nat Neurosci.
2012;15:816–8. Article CAS PubMed PubMed Central Google Scholar * Saga Y, Richard A, Sgambato-Faure V, Hoshi E, Tobler PN, Tremblay L. Ventral pallidum encodes contextual information
and controls aversive behaviors. Cereb Cortex. 2017;27:2528–43. PubMed Google Scholar * Grillon C, Baas JMP, Cornwell B, Johnson L. Context conditioning and behavioral avoidance in a
virtual reality environment: effect of predictability. Biol Psychiatry. 2006;60:752–9. Article PubMed Google Scholar * Krypotos A-M, Effting M, Arnaudova I, Kindt M, Beckers T. Avoided by
association: acquisition, extinction, and renewal of avoidance tendencies toward conditioned fear stimuli. Clin Psychol Sci. 2014;2:336–43. Article Google Scholar * Lau JYF, Viding EM.
Anxiety-related biases in children’s avoidant responses to a masked angry face. Behav Res Ther. 2007;45:1639–45. Article PubMed Google Scholar * Jafari-Sabet M. NMDA receptor antagonists
antagonize the facilitatory effects of post-training intra-basolateral amygdala NMDA and physostigmine on passive avoidance learning. Eur J Pharm. 2006;529:122–8. Article CAS Google
Scholar * Oler JA, Fox AS, Shackman AJ, Kalin NH. The central nucleus of the amygdala is a critical substrate for individual differences in anxiety. In: Amaral DG, editor. Living without an
amygdala, (The Guilford Press: 2016. p. 218–51, vol. 430, New York. * Choi J-S, Kim JJ. Amygdala regulates risk of predation in rats foraging in a dynamic fear environment. Proc Natl Acad
Sci USA. 2010;107:21773–7. Article CAS PubMed PubMed Central Google Scholar * Kenwood MM, Kalin NH. Nonhuman primate models to explore mechanisms underlying early-life temperamental
anxiety. Biol Psychiatry. 2021;89:659–71. Article CAS PubMed Google Scholar * Kyriazi P, Headley DB, Pare D. Multi-dimensional coding by basolateral amygdala neurons. Neuron.
2018;99:1315–28 e5. Article CAS PubMed PubMed Central Google Scholar * Treanor M, Barry TJ. Treatment of avoidance behavior as an adjunct to exposure therapy: Insights from modern
learning theory. Behav Res Ther. 2017;96:30–36. Article PubMed Google Scholar * Smith BM, Smith GS, Dymond S. Relapse of anxiety‐related fear and avoidance: Conceptual analysis of
treatment with acceptance and commitment therapy. Jrnl Exp Anal Behav. 2020;113:87–104. Article Google Scholar * Bravo-Rivera C, Roman-Ortiz C, Montesinos-Cartagena M, Quirk GJ. Persistent
active avoidance correlates with activity in prelimbic cortex and ventral striatum. Front Behav Neurosci. 2015;9:184. Article PubMed PubMed Central Google Scholar * Martinez-Rivera FJ,
Bravo-Rivera C, Velazquez-Diaz CD, Montesinos-Cartagena M, Quirk GJ. Prefrontal circuits signaling active avoidance retrieval and extinction. Psychopharmacology 2019;236:399–406. Article
CAS PubMed Google Scholar * Gillan CM, Morein-Zamir S, Urcelay GP, Sule A, Voon V, Apergis-Schoute AM, et al. Enhanced avoidance habits in obsessive-compulsive disorder. Biol Psychiatry.
2014;75:631–8. Article PubMed PubMed Central Google Scholar * Martínez-Rivera FJ, Sánchez-Navarro MJ, Huertas-Pérez CI, Greenberg BD, Rasmussen SA, Quirk GJ. Prolonged avoidance training
exacerbates OCD-like behaviors in a rodent model. Transl Psychiatry. 2020;10:212. Article PubMed PubMed Central Google Scholar * Rodriguez-Romaguera J, Greenberg BD, Rasmussen SA, Quirk
GJ. An avoidance-based rodent model of exposure with response prevention therapy for obsessive-compulsive disorder. Biol Psychiatry. 2016;80:534–40. Article PubMed PubMed Central Google
Scholar * Gremel CM, Costa RM. Orbitofrontal and striatal circuits dynamically encode the shift between goal-directed and habitual actions. Nat Commun. 2013;4:2264. Article PubMed Google
Scholar * Graybiel AM, Rauch SL. Toward a neurobiology of obsessive-compulsive disorder. Neuron. 2000;28:343–7. Article CAS PubMed Google Scholar * Gillan CM, Apergis-Schoute AM,
Morein-Zamir S, Urcelay GP, Sule A, Fineberg NA, et al. Functional neuroimaging of avoidance habits in obsessive-compulsive disorder. Am J Psychiatry. 2015;172:284–93. Article PubMed
Google Scholar * Norbury A, Robbins TW, Seymour B. Value generalization in human avoidance learning. Elife. 2018;7:e34779. * Ciocchi S, Herry C, Grenier F, Wolff SBE, Letzkus JJ, Vlachos I,
et al. Encoding of conditioned fear in central amygdala inhibitory circuits. Nature. 2010;468:277–82. Article CAS PubMed Google Scholar * Likhtik E, Stujenske JM, Topiwala MA, Harris
AZ, Gordon JA. Prefrontal entrainment of amygdala activity signals safety in learned fear and innate anxiety. Nat Neurosci. 2014;17:106–13. Article CAS PubMed Google Scholar * Lommen
MJJ, Engelhard IM, van den Hout MA. Neuroticism and avoidance of ambiguous stimuli: Better safe than sorry? Pers Individ Dif. 2010;49:1001–6. Article Google Scholar * Boyle S, Roche B,
Dymond S, Hermans D. Generalisation of fear and avoidance along a semantic continuum. Cogn Emot. 2016;30:340–52. Article PubMed Google Scholar * Dymond S, Schlund MW, Roche B, Whelan R.
The spread of fear: symbolic generalization mediates graded threat-avoidance in specific phobia. Q J Exp Psychol. 2014;67:247–59. Article Google Scholar * van Meurs B, Wiggert N, Wicker I,
Lissek S. Maladaptive behavioral consequences of conditioned fear-generalization: a pronounced, yet sparsely studied, feature of anxiety pathology. Behav Res Ther. 2014;57:29–37. Article
PubMed PubMed Central Google Scholar * Sheynin J, Moustafa AA, Beck KD, Servatius RJ, Myers CE. Testing the role of reward and punishment sensitivity in avoidance behavior: A
computational modeling approach. Behav Brain Res. 2015;283:121–38. Article PubMed PubMed Central Google Scholar * Bravo-Rivera H, Rubio Arzola P, Caban-Murillo A, Vélez-Avilés AN,
Ayala-Rosario SN, Quirk GJ. Characterizing different strategies for resolving approach-avoidance conflict. Front Neurosci. 2021;15:608922. Article PubMed PubMed Central Google Scholar *
Fanselow MS, Lester LS, Helmstetter FJ. Changes in feeding and foraging patterns as an antipredator defensive strategy: a laboratory simulation using aversive stimulation in a closed
economy. J Exp Anal Behav. 1988;50:361–74. Article CAS PubMed PubMed Central Google Scholar * Schuessler BP, Zambetti PR, Kukuoka KM, Kim EJ, Kim JJ. The risky closed economy: a
holistic, longitudinal approach to studying fear and anxiety in rodents. Front Behav Neurosci. 2020;14:594568. Article PubMed PubMed Central Google Scholar * Kim E, Kim EJ, Yeh R, Shin
M, Bobman J, Krasne FB, et al. Amygdaloid and non-amygdaloid fear both influence avoidance of risky foraging in hungry rats. Proc Biol Sci. 2014;281:20133357. * Pittig A, Scherbaum S. Costly
avoidance in anxious individuals: elevated threat avoidance in anxious individuals under high, but not low competing rewards. J Behav Ther Exp Psychiatry. 2020;66:101524. Article PubMed
Google Scholar * Korn CW, Vunder J, Miró J, Fuentemilla L, Hurlemann R, Bach DR. Amygdala lesions reduce anxiety-like behavior in a human benzodiazepine-sensitive approach–avoidance
conflict test. Biol Psychiatry. 2017;82:522–31. Article PubMed PubMed Central Google Scholar * Abivardi A, Khemka S, Bach DR. Hippocampal representation of threat features and behavior
in a human approach–avoidance conflict anxiety task. J Neurosci. 2020;40:6748–58. Article CAS PubMed PubMed Central Google Scholar * Aupperle RL, Melrose AJ, Francisco A, Paulus MP,
Stein MB. Neural substrates of approach-avoidance conflict decision-making. Hum Brain Mapp. 2015;36:449–62. Article PubMed Google Scholar * Machado CJ, Kazama AM, Bachevalier J. Impact of
amygdala, orbital frontal, or hippocampal lesions on threat avoidance and emotional reactivity in nonhuman primates. Emotion. 2009;9:147–63. Article PubMed PubMed Central Google Scholar
* Saga Y, Ruff CC, Tremblay L. Disturbance of approach‐avoidance behaviors in non‐human primates by stimulation of the limbic territories of basal ganglia and anterior insula. Eur J
Neurosci. 2019;49:687-700. * La-Vu M, Tobias BC, Schuette PJ, Adhikari A. To approach or avoid: an introductory overview of the study of anxiety using rodent assays. Front Behav Neurosci.
2020;14:145. Article PubMed PubMed Central Google Scholar * Carobrez AP, Bertoglio LJ. Ethological and temporal analyses of anxiety-like behavior: the elevated plus-maze model 20 years
on. Neurosci Biobehav Rev. 2005;29:1193–205. Article CAS PubMed Google Scholar * Friedman A, Homma D, Gibb LG, Amemori K-I, Rubin SJ, Hood AS, et al. A corticostriatal path targeting
striosomes controls decision-making under conflict. Cell. 2015;161:1320–33. Article CAS PubMed PubMed Central Google Scholar * Amemori K-I, Graybiel AM. Localized microstimulation of
primate pregenual cingulate cortex induces negative decision-making. Nat Neurosci. 2012;15:776–85. Article CAS PubMed PubMed Central Google Scholar * Loewke AC, Minerva AR, Nelson AB,
Kreitzer AC, Gunaydin LA. Frontostriatal projections regulate innate avoidance behavior. J Neurosci. 2021;41:5487–501. Article CAS PubMed PubMed Central Google Scholar * Lebowitz ER,
François B. Using motion tracking to measure avoidance in children and adults: psychometric properties, associations with clinical characteristics, and treatment-related change. Behav Ther.
2018;49:853–65. Article PubMed PubMed Central Google Scholar * Kallai J, Karádi K, Bereczkei T, Rózsa S, Jacobs WJ, Nadel L. Spatial exploration behaviour in an extended labyrinth in
patients with panic disorder and agoraphobia. Psychiatry Res. 2007;149:223–30. Article PubMed Google Scholar * Walz N, Mühlberger A, Pauli P. A human open field test reveals thigmotaxis
related to agoraphobic fear. Biol Psychiatry. 2016;80:390–7. Article PubMed Google Scholar * Biedermann SV, Biedermann DG, Wenzlaff F, Kurjak T, Nouri S, Auer MK, et al. An elevated
plus-maze in mixed reality for studying human anxiety-related behavior. BMC Biol. 2017;15:125. Article PubMed PubMed Central Google Scholar * Freeman D, Reeve S, Robinson A, Ehlers A,
Clark D, Spanlang B, et al. Virtual reality in the assessment, understanding, and treatment of mental health disorders. Psychol Med. 2017;47:2393–2400. Article CAS PubMed PubMed Central
Google Scholar * Onnela JP. Opportunities and challenges in the collection and analysis of digital phenotyping data. Neuropsychopharmacology. 2021;46:45–54. Article PubMed Google Scholar
* Stephan KE, Iglesias S, Heinzle J, Diaconescu AO. Translational perspectives for computational neuroimaging. Neuron. 2015;87:716–32. Article CAS PubMed Google Scholar * Mitchell AS,
Thiele A, Petkov CI, Roberts A, Robbins TW, Schultz W, et al. Continued need for non-human primate neuroscience research. Curr Biol. 2018;28:R1186–R1187. Article CAS PubMed PubMed Central
Google Scholar * Ironside M, Amemori K-I, McGrath CL, Pedersen ML, Kang MS, Amemori S, et al. Approach-avoidance conflict in major depressive disorder: congruent neural findings in humans
and nonhuman primates. Biol Psychiatry. 2020;87:399–408. Article PubMed Google Scholar * McDermott TJ, Kirlic N, Akeman E, Touthang J, Clausen AN, Kuplicki R, et al. Test-retest
reliability of approach-avoidance conflict decision-making during functional magnetic resonance imaging in healthy adults. Hum Brain Mapp. 2021;42:2347–61. Article PubMed PubMed Central
Google Scholar * Vervliet B, Raes F. Criteria of validity in experimental psychopathology: application to models of anxiety and depression. Psychol Med. 2013;43:2241–4. Article CAS PubMed
Google Scholar * Olatunji BO, Cisler JM, Deacon BJ. Efficacy of cognitive behavioral therapy for anxiety disorders: a review of meta-analytic findings. Psychiatr Clin North Am.
2010;33:557–77. Article PubMed Google Scholar * Craske MG, Treanor M, Conway CC, Zbozinek T, Vervliet B. Maximizing exposure therapy: an inhibitory learning approach. Behav Res Ther.
2014;58:10–23. Article PubMed PubMed Central Google Scholar * Arch JJ, Craske MG. Acceptance and commitment therapy and cognitive behavioral therapy for anxiety disorders: different
treatments, similar mechanisms? Clin Psychol: Sci Pract. 2008;15:263–79. Google Scholar Download references ACKNOWLEDGEMENTS We would like to thank Dr. Tom Neylan for feedback on the
manuscript. Figures were created with BioRender.com. FUNDING LAG is funded by the Chan Zuckerberg Biohub and UCSF Program for Breakthrough Biomedical Research. While conducting this work,
TMB was funded by the Brain and Behavioral Research Foundation (Grant #27233) and National Institute of Mental Health (K23MH113708). TMB now receives salary from Big Health Inc, for work
unrelated to this manuscript. LAG reports no financial relationships with commercial interests. AUTHOR INFORMATION AUTHORS AND AFFILIATIONS * Department of Psychiatry and Behavioral
Sciences, Stanford University School of Medicine, Stanford, CA, 94305, USA Tali M. Ball * Department of Psychiatry and Behavioral Sciences, University of California San Francisco, San
Francisco, CA, 94143, USA Lisa A. Gunaydin * Kavli Institute for Fundamental Neuroscience, University of California San Francisco, San Francisco, CA, 94143, USA Lisa A. Gunaydin Authors *
Tali M. Ball View author publications You can also search for this author inPubMed Google Scholar * Lisa A. Gunaydin View author publications You can also search for this author inPubMed
Google Scholar CONTRIBUTIONS TMB and LAG contributed equally to the conception, design, and writing of the work. CORRESPONDING AUTHORS Correspondence to Tali M. Ball or Lisa A. Gunaydin.
ADDITIONAL INFORMATION PUBLISHER’S NOTE Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations. RIGHTS AND PERMISSIONS OPEN
ACCESS This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format,
as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third
party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the
article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright
holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/. Reprints and permissions ABOUT THIS ARTICLE CITE THIS ARTICLE Ball, T.M., Gunaydin, L.A. Measuring
maladaptive avoidance: from animal models to clinical anxiety. _Neuropsychopharmacol._ 47, 978–986 (2022). https://doi.org/10.1038/s41386-021-01263-4 Download citation * Received: 16 June
2021 * Revised: 04 November 2021 * Accepted: 22 December 2021 * Published: 15 January 2022 * Issue Date: April 2022 * DOI: https://doi.org/10.1038/s41386-021-01263-4 SHARE THIS ARTICLE
Anyone you share the following link with will be able to read this content: Get shareable link Sorry, a shareable link is not currently available for this article. Copy to clipboard Provided
by the Springer Nature SharedIt content-sharing initiative