Constitutive activation of two-component systems reveals regulatory network interactions in streptococcus agalactiae
- Select a language for the TTS:
- UK English Female
- UK English Male
- US English Female
- US English Male
- Australian Female
- Australian Male
- Language selected: (auto detect) - EN
Play all audios:

ABSTRACT Bacterial two-component systems (TCSs) are signaling modules that control physiology, adaptation, and host interactions. A typical TCS consists of a histidine kinase (HK) that
activates a response regulator via phosphorylation in response to environmental signals. Here, we systematically test the effect of inactivating the conserved phosphatase activity of HKs to
activate TCS signaling pathways. Transcriptome analyses of 14 HK mutants in _Streptococcus agalactiae_, the leading cause of neonatal meningitis, validate the conserved HK phosphatase
mechanism and its role in the inhibition of TCS activity in vivo. Constitutive TCS activation, independent of environmental signals, enables high-resolution mapping of the regulons for
several TCSs (e.g., SaeRS, BceRS, VncRS, DltRS, HK11030, HK02290) and reveals the functional diversity of TCS signaling pathways, ranging from highly specialized to interconnected global
regulatory networks. Targeted analysis shows that the SaeRS-regulated PbsP adhesin acts as a signaling molecule to activate CovRS signaling, thereby linking the major regulators of
host-pathogen interactions. Furthermore, constitutive BceRS activation reveals drug-independent activity, suggesting a role in cell envelope homeostasis beyond antimicrobial resistance. This
study highlights the versatility of constitutive TCS activation, via phosphatase-deficient HKs, to uncover regulatory networks and biological processes. SIMILAR CONTENT BEING VIEWED BY
OTHERS AN _E. COLI_ DISPLAY METHOD FOR CHARACTERIZATION OF PEPTIDE–SENSOR KINASE INTERACTIONS Article 08 December 2022 HIGHLY-CONSERVED REGULATORY ACTIVITY OF THE ANR FAMILY IN THE VIRULENCE
OF DIARRHEAGENIC BACTERIA THROUGH INTERACTION WITH MASTER AND GLOBAL REGULATORS Article Open access 29 April 2023 IMPACT OF _ABAI_ MUTATION ON VIRULENCE, BIOFILM DEVELOPMENT, AND ANTIBIOTIC
SUSCEPTIBILITY IN _ACINETOBACTER BAUMANNII_ Article Open access 14 September 2024 INTRODUCTION Two-component systems (TCSs) are one of the main bacterial signalling mechanisms. In their
simplest form, an environmental signal activates a histidine kinase (HK), which phosphorylates a cognate response regulator (RR), leading to the transcription of specific genes that mediate
the cellular response to the stimuli1. Actually, TCSs are sophisticated molecular machinery with buffering and insulating mechanisms that dynamically control specific or global cellular
responses2,3,4. Considerable effort has been made to define TCS regulatory networks in both model and pathogenic species, including by comprehensive analysis5,6. Although knowledge gained in
one species can provide information about homologous systems, TCS are characterised by their diversity, plasticity and evolvability7,8. This prevents global inferences even between closely
related species9,10. This evolution of regulatory networks is sustained by several mechanisms, including mutations, horizontal gene transfer, duplication followed by neofunctionalization,
and rewiring that shapes adaptation and speciation11,12,13. Functional, evolutionary, and system analyses require characterising individual signalling pathways and integrating them into the
cellular regulatory network. Traditionally, regulons are characterised using inactivated TCS mutants. One common pitfall is that TCSs are not active until the specific, but usually unknown,
stimulus is provided. Current approaches to overcome signal requirements are based on phosphomimetic mutation of the RR14,15 and profiling of protein-DNA interaction16,17,18. An alternative
approach exploits the distinct HK enzymatic activities. The HK cytoplasmic core called the transmitter module, is composed of the DHp (Dimerisation and Histidine phosphotransfer) and CA
(catalytic and ATP-binding) domains19,20,21. The two domains are dynamically structured in specific conformations that catalyse three distinct reactions: autophosphorylation of a conserved
histidine residue in the DHp domain, phosphotransfer to a conserved aspartate on the RR, and RR dephosphorylation. Pioneering studies have identified mutations abolishing the HK phosphatase
activity leading to increased RR phosphorylation and signalling pathway activation22,23,24. The importance of HK phosphatase activity in vivo has been initially debated, especially when
considering the lability of RR phosphorylation and spontaneous dephosphorylation rate25. Nowadays, phosphatase activity is recognised as essential for the dynamics of the response and to
ensure that the RR is activated by the cognate HK only26,27. Co-evolving residues and HKs conformational rearrangements ensure the specificity and directionality of enzymatic
reactions28,29,30. While the activation mechanism involving the conserved histidine residue is fundamentally conserved among HKs, the phosphatase mechanism has remained more elusive due to
variations in the DHp domain31. Then, a seminal study proposed a conserved phosphatase mechanism for the two main HisKA and HisKA_3 families, identifying conserved motifs and specific
catalytic residues needed for the correct positioning of nucleophilic attack31,32. Substitution of the catalytic residues abolishes the phosphatase activity without impacting the autokinase
and phosphotransfer activities, resulting in increased RR phosphorylation and pathway activation for the individual HKs reported to date32,33,34,35,36,37. This study aims to systematically
test the proposed conserved mechanism of phosphatase activity, the in vivo effect of phosphatase-deficient HK, and the activation of the regulatory network in all HisKA and HisKA_3 systems
in a bacterium. We focused on _Streptococcus agalactiae_ (Group B _Streptococcus_, GBS), a pathobiont that is commensal in adults but pathogenic during pregnancy and in neonates, for whom it
is the leading cause of invasive infections38,39. We report that targeting HK phosphatase activity provides high-resolution views of signalling pathways for most TCSs independently from
environmental signals. In addition, regulatory network activation resolves the connectivity between TCSs involved in host-pathogen interactions and reveals the physiological function of a
TCS involved in antimicrobial resistance. This systematic analysis argues for the widespread adoption of this gain-of-function approach to decipher TCSs signalling in genetically manipulable
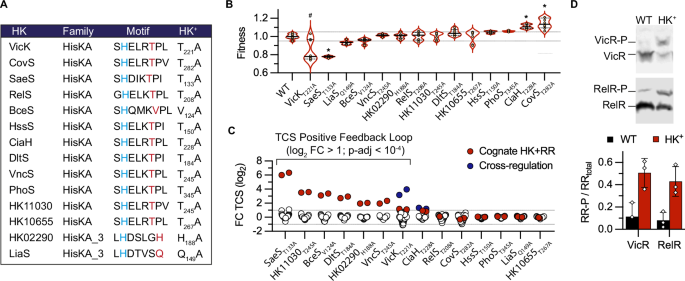
species. RESULTS THE HK+ COLLECTION TARGETS THE PHOSPHATASE ACTIVITY OF HISTIDINE KINASE We undertook a genetic approach to systematically test the hypothesis of a conserved
dephosphorylation mechanism in the two major HK families32. The genome of the BM110 strain belonging to the hypervirulent clonal complex 17 (CC–17) encodes 20 HKs40, among which 12 and 2
have a HisKA and HisKA_3 DHp domain, respectively (Supplementary Data 1). Their H-box motif always contains the conserved phospho-acceptor histidine, immediately followed by the predicted
phosphatase motif (Fig. 1A). Eleven of the twelve HisKA proteins have the E/DxxT/N motif with a putative threonine catalytic residue, while the remaining HisKA protein (BceS) has a divergent
sequence composition (QMKV) with a valine at the predicted catalytic position (Fig. 1A). The two HisKA_3 proteins have the DxxxQ/H motif with the predicted glutamine or histidine catalytic
residue (Fig. 1A). The 14 HKs encoding genes are organised in operon with their cognate response regulator (RR) belonging to the OmpR (with HisKA) or LuxR (with HisKA_3) family, but one
system is not functional (HK10655-RR10650fs) due to a pseudogenization of the RR in the CC-17 hypervirulent GBS lineage (Supplementary Data 1). We generated 14 strains, called the HK+
collection, with an alanine substitution of the predicted phosphatase catalytic residue (Fig. 1A). Whole-genome sequencing confirmed the chromosomal substitution of targeted base pairs and
the absence of secondary mutations in 11 out of the 14 HK+ strains. In the three remaining HK+ (CovST282A, VicKT221A, and RelST208A), we sequenced independent mutants and selected one with a
single secondary mutation (Supplementary Data 2). Notably, the selected VicKT221A mutant possesses a non-synonymous polymorphism in the glutamine transporter GlnPQ, which we cannot dismiss
as a potential compensatory mutation. Four independently constructed VicKT221A mutants exhibit putative compensatory mutations (Supplementary Data 2), a phenomenon commonly observed in
mutants within the homologous WalRK system, which is essential for cell wall remodelling during growth and division41,42,43. Individual growth curves show a significant effect ( | F | >
0.1, Mann Whitney test _p_ < 10−4) of the HK+ mutation for four mutants (Fig. 1B). The CovST282A and CiaHT228A have a reproducible fitness advantage compared to the WT strain, while the
VicKT221A and SaeST133A have fitness defect. Notably, the slow-growing VicKT221A mutant is unstable and gives rise to faster-growing cultures, likely due to additional mutations, while the
SaeST133A mutant exhibits a density-dependent phenotype characterised by a decreasing growth rate in the exponential phase and a lower final OD (Supplementary Fig. 1). In addition, two
mutants have increased antibiotic susceptibilities: the VicKT221A mutant against beta-lactams, in agreement with a conserved function in cell wall metabolism, and the RelS mutant against
fosfomycin (Supplementary Data 3). HK+ ACTIVATE POSITIVE FEEDBACK LOOPS To test TCS activation, we first relied on positive feedback loops. This autoregulation is often observed through
direct transcriptional activation of the TCS operon by the activated RR2. We therefore analysed the transcription of all HKs and RRs encoding genes (_n_ = 41, including non-HisKA and HisKA_3
TCSs and an orphan RR) in each HK+ mutant by RNA-sequencing from cultures grown in a standardised condition (THY: Todd Hewitt supplemented with 1% Yeast extract, pH 7.4, 37 °C, exponential
growth phase OD600 = 0.5). A positive feedback loop, indicated by a significant fold change greater than two relative to the WT strain (log2 FC > 1; p-adj < 10−4) for the HK and RR
genes, is observed in seven HK+ mutants (Fig. 1C). Furthermore, two TCSs are significantly regulated in an unrelated HK+ mutant: the HK11050-RR11055 system, which does not contain a HisKA
and HisKA_3 domain, in the VicKT221A mutant and the RelRS system, which is not positively auto-regulated, in the CiaHT228A mutant (Fig. 1C). As an independent approach to test TCS
activation, we introduced in each mutant a vector expressing an epitope-tagged copy of the cognate regulator. For two mutants (VicKT221A and RelST208A), an increased level of phosphorylation
of the ectopically expressed regulator is detected in the HK+ mutant compared to the WT strain after Phos-Tag electrophoresis and western analysis with anti-FLAG antibodies (Fig. 1D).
However, due to competition between WT and epitope-tagged regulators and variability in the stability of phosphorylated aspartate, no conclusions could be drawn for most regulators. This
highlights the need to quantify the level of phosphorylation using specific antibodies directed against each native RR. Overall, by considering epitope-tagged RR activation by
phosphorylation and positive feedback loops, the majority (8/14) of HK+ mutations appear to activate the corresponding TCS signalling pathway. THE ACTIVATED GENE REGULATORY NETWORK To
characterise the activated pathways, we analysed the RNA-seq profiles of each HK+ mutant grown under standardised conditions, independent of specific environmental cues (i.e., exponential
phase in THY pH 7.4 at 37 °C). Statistical analysis of differentially expressed genes (DEGs: Supplementary Data 4) grouped the HK+ mutants into three main categories based on the adjusted
_p_-values. Six HK+ mutants (HK11030T245A, VncST245A, SaeST133A, BceSV124A, HK02290H188A, and DltST184A) show DEGs associated with striking statistical significance (p-adj < 10−250),
revealing the activated regulons with high resolution (Fig. 2A and Supplementary Fig. 2). Four additional mutants (RelST208A, CiaHT228A, VicKT221A, and CovST282A) show DEGs with lower
statistical significance (p-adj > 10−150), suggesting complex regulons or intermediate TCS activation (Supplementary Fig. 2). The remaining four mutants (HssST150A, LiaSQ149A, PhoST345A,
and the HK10655T267A with a frameshifted RR) gave no or low significant signals (p-adj > 10−10) (Supplementary Fig. 2). Since most RRs are transcriptional activators, we focused the
analysis on activated genes. By applying strict thresholds (FC > 3, p-adj < 10−4) for normalisation between samples and excluding genes with very low read counts in all samples and
genes localised in mobile genetic elements, 219 genes (11.9% of the 1838 genes analysed) are transcriptionally activated in at least one HK+ mutant (Supplementary Data 4E). Transcriptional
activation can be up to 8000-fold, with an average fold change of 61.6-fold and an uneven distribution between HK+ mutants (Fig. 2B). The number of activated genes ranges from 3
(HK11030T245A) to 139 (VicKT221A) (Fig. 2B and Supplementary Data 4F). Five regulatory systems activate a specific genetic programme, four of them (HK11030T245A, HK02290H188A, VncST245A,
DltST184A) positively regulating a single functional genetic module composed of their own operon and at least one additional gene involved in the cellular response localised into, or
adjacent to, the TCS operon (Fig. 2C), and one system (CiaHT228A) coordinating the activation of at least six independent loci (Supplementary Fig. 3). Four additional TCSs activate specific
genes but share 1 to 3 activated genes with the VicKT221A mutant (Fig. 2D). One of these connected systems (HssST150A) is specialised in haem detoxification via the transcriptional
activation of the _hrtBA_ genes encoding a specific ABC transporter44, which is similarly activated in the VicKT221A mutant. The three additional connected systems activate several loci
involved in host-pathogen interaction (SaeST133A: adhesins and secreted proteins), drug resistance (BceSV124A: transporters and peptidase), or nucleotide metabolism (RelST208A: de novo
purine synthesis and ectonucleotidases), with (SaeST133A, BceSV124A) or without (RelST208A) a positive feedback loop (Fig. 2D and Supplementary Fig. 3). POSITIVE AND NEGATIVE INTERACTION
BETWEEN TCS SYSTEMS Overall, the HK+ mutation activates the signalling pathway for 10 out of 12 TCSs (Fig. 2D), excluding the CovST282A repressing system analysed separately and the negative
control HK10655T267A with a frameshifted RR. Notably, each HK+ mutant is associated with the activation of specific genes, except the global regulator VicRK (Fig. 2D and Supplementary Data
4E). As expected, the VicKT221A regulon included several operons involved in cell wall metabolism (Supplementary Fig. 3). However, constitutive activation of VicK probably leads to the
activation of related stress and cell-wall signalling pathways. To identify relationships between TCS pathways involved in related processes, we analysed the 219 genes activated in at least
one HK+ mutant for their expression in the whole RNA-seq dataset. This analysis confirmed the partial activation of SaeRS signalling in the VicKT221A mutant (a shared CovR connection, in
fact, see specific section below) sustained by genes with 1 < FC < 3 and significant but higher p-adj value compared to the SaeST133A activated system (Supplementary Data 4H).
Similarly, by considering significantly regulated genes with lower thresholds (1 < | FC | < 3, p-adj< 0.05), significant positive or antagonistic interactions were detected between
signalling pathways (e.g., DltS activating CiaH and VicK, CiaH antagonising RelS, HK02290 antagonising HK11030). Finally, relaxing the thresholds also reveals the first five genes of the
_phoRS_ operon as the most and only significantly up-regulated genes (1,5 < FC < 1,75; 7.10−3 < p-adj< 10−5) in the PhoST345A mutant (Supplementary Data 4D), suggesting a
conserved mechanism of phosphatase activity but an inefficient activation of the PhoR regulator in the corresponding HK+ mutant. ACTIVATION OF THE GLOBAL REPRESSOR OF VIRULENCE COVRS The
CovRS system is the major regulator of virulence in GBS and, in contrast to canonical TCS, acts as a global repressor of gene transcription45. Analysis of RNA-seq to identify negative
regulation using similar thresholds (− 3 > FC, p-adj < 10−4) revealed the repression of 32 genes in the whole dataset (Supplementary Data 4G). Almost all repressed genes are regulated
by the two global regulators, VicK (17 genes) and CovS (14 genes). Notably, the most highly repressed gene (22-fold in VicKT221A) encodes a D-L endopeptidase, highlighting the conservation
of negative regulation of cell wall hydrolases between WalRK homologous systems41. On the other hand, a comparative analysis of repressed genes in CovST282A with the known CovR regulon shows
the limitations of the HK+ approach for characterising the CovRS system. Indeed, only 5 out of 14 repressed genes in the CovST282A mutant belong to the CovR regulon of 153 genes previously
determined with loss of function mutants45. This difference can be attributed to another specific feature of the CovRS system, which is active in the absence of an environmental signal45.
The CovST282A transcriptome, therefore, suggests that CovR over-activation does not translate into increased repression of targeted genes. To further analyse the CovRS system, we compared
the transcriptome of the activated CovST282A mutant with that of the inactivated CovRD53A mutant, which cannot be phosphorylated by CovS, and also included our genome-wide CovR binding
analysis done by ChIP-seq45. Side-by-side transcriptome comparison showed that the inactivation of CovR activates the signalling pathway repressed by the active CovRS system, without a
general inverse relationship in the CovST282A mutant (Fig. 3A). Nevertheless, a significant inverse correlation between the CovST282A and CovRD53A transcriptomes is observed for genes that
do not belong to the direct CovR regulon (Fig. 3B), suggesting that overactivation of CovR primarily increases binding to co-regulated promoters and low-affinity binding sites45. Since the
inactivation of the CovRS repressor system is more informative than its over-activation, we included the CovRD53A transcriptome in the HK+ dataset. The global gene network reveals
connections between CovR-repressed genes and SaeST133A, VicKT221A, or BceSV124A activated genes (Fig. 2D). Notably, SaeST133A is highly connected with the direct CovR-repressed genes, while
BceSV124A activates only three CovR indirectly regulated genes. Functional assays using pigmented beta-haemolysin/cytolysin (ß-h/c) production as a natural reporter of CovR activity first
confirmed the non-pigmented and non-haemolytic phenotypes of the CovST282A mutant (Fig. 3C), in agreement with CovR directly repressing the _cyl_ operon encoding the ß-h/c synthesis and
export machineries45,46. The phenotype of six additional HK+ mutants are different from the WT strain on selective media, either increasing (SaeST133A, VicKT221A, and RelST208A) or
decreasing (BceSV124A, CiaHT228A, and LiaSQ149A) pigmentation and/or haemolytic activity (Fig. 3C). However, the absence of correlation between the transcription of the _cyl_ operon and the
pigmentation/haemolytic phenotypes in several HK+ mutants (Fig. 3C) suggests that ß-h/c activity depends on post-transcriptional regulatory mechanisms in addition to CovR regulation of the
_cyl_ operon. This is potentially the case in mutants with altered cell surface composition, where the toxin interaction occurs47,48. THE PBSP ADHESIN CONNECTS SAERS AND COVRS SIGNALLING We
sought to decipher the connection between the CovRS and SaeRS systems, two main regulators of host-pathogen interactions. Published transcriptomes with _saeRS_ deletion mutants define a
large regulon of 400–600 genes depending on growth conditions49. In contrast, analysis of the SaeST133A transcriptome reveals the highly significant (60 < FC < 8000-fold, p-adj <
10−250) activation of four genes only, along with a partial activation of the CovR regulon (Fig. 4A). We confirmed the stratification of the SaeST133A differentially regulated genes by
RT-qPCR, validating 3 groups: the _pbsP_ and _bvaP_ genes, the _saeRS_ operon, and the CovR-regulated genes represented by the directly repressed genes _cylE_ and _hvgA_ (Fig. 4B). Intrigued
by the almost 50-fold difference between _pbsP_ and _saeRS_ up-regulation, we analysed the genomic locus in detail. The 112 bp _pbsP_-_saeRS_ intergenic region contains a P_saeR_ promoter
but no canonical transcriptional terminator. The integration of such a terminator precisely after the _pbsP_ stop codon in the SaeST133A mutant abolishes _saeRS_ overexpression while having
no impact on other activated genes (Fig. 4B). Quantification of promoter activities using _ß-_galactosidase reporters confirms a similar activity of P_saeR_ in the WT and in SaeST133A
mutant, and the strong activation of P_pbsP_ upon activation of the SaeRS system (Fig. 4C). This shows an indirect positive feedback loop of the _saeRS_ operon, which is transcribed by its
constitutive promoter and regulated by _pbsP_ termination readthrough. Interestingly, the basal level of _saeRS_ transcription in SaeST133A with the _pbsP_ terminator is sufficient to fully
activate _pbsP_ and _bvaP_ (Fig. 4B), implying that the indirect feedback loop may be physiologically relevant only for controlling the kinetics of the signal-dependent response, but not its
amplitude. We next analysed the connection between SaeRS and CovRS signalling. The activation of CovR-regulated genes in the SaeST133A mutant is intermediate when compared to the CovRD53A
mutant (Figs. 3A, 4A). One hypothesis could be a competitive binding between SaeR and CovR, but it is unlikely that all binding sites will allow both SaeR-activation and CovR-repression. As
an alternative, we hypothesised that the two genes specifically regulated in the SaeST133A mutant, encoding the PbsP cell-wall anchored adhesin50,51 and the BvaP secreted protein52, could be
involved in the activation of the CovR regulon. Indeed, the deletion of _pbsP_, but not of _bvaP_, in the SaeST133A mutant, restores a WT haemolytic activity (Fig. 4D). In agreement with
the phenotypes, the deletion of _pbsP_ in the SaeST133A mutant restores a WT level of the CovR-regulated genes _cylE_ and _hvgA_, while the _saeRS_ and _bvaP_ genes are still similarly
up-regulated (Fig. 4E). After cleavage by the enzyme sortase A and anchoring to the cell wall, the remaining carboxy-terminal domain of an LPxTG adhesin can act as a signalling molecule by
interacting with the transmembrane domain of a specific HK, as demonstrated in _Streptococcus gordonii_53. We, therefore, considered this C-peptide mechanism and constructed mutants
expressing truncated PbsP variants in the SaeST133A mutant. In-frame deletion of the PbsP C-peptide (e.g., 108 bp deletion including the LPxTG cell-wall anchoring motif until the penultimate
codon) has no effect on the induction of the CovR-regulated haemolytic activity (Fig. 4D). In contrast, in-frame deletion of the PbsP adhesin domain (1239 bp deletion leaving the signal
peptide and the LPxTG cell wall anchoring motif intact) restores the haemolytic activity of the SaeST133A mutant to WT level (Fig. 4D). Furthermore, the growth defect of the SaeST133A
mutant, which is similar to the growth defect of the CovRD53A mutant, is suppressed by deletion of _pbsP_ or of the adhesin part of _pbsP_ (Supplementary Fig. 4). Thus, the PbsP adhesin
domain triggers CovR signalling either by interacting with CovS or co-regulatory proteins54 or by inducing surface perturbations specifically sensed by the CovRS system (Fig. 4F).
DRUG-INDEPENDENT ACTIVITY OF THE BCERS THREE-COMPONENT SYSTEM We next sought to decipher the function of the BceRS system, which, upon activation, shows hypo-pigmented and haemolytic
phenotypes that are independent of CovRS regulation of the _cyl_ operon (Fig. 3C). The BceRS system belongs to a conserved TCS family that relies on a transporter to sense and transmit
environmental signals to the HK55. The transcriptome of the BceSV124A mutant reveals a 9-gene regulon, including the _bceRS_ operon and adjacent genes (Fig. 5A and Supplementary Figs. 2 and
3). Further validation by RT-qPCR confirmed the 10- to 1000-fold activation of the regulon in the absence of drugs in the BceSV124A mutant, as well as the absence of significant
transcriptional changes of the CovR-regulated genes (Fig. 5B). As expected, mutation of the BceR regulator to a non-phosphorylated form (BceRD55A) abolishes the activation of the signalling
pathway in the BceSV124A mutant (Fig. 5B). Interestingly, deletion of the transporter/sensor (∆_bceAB_) in the BceSV124A mutant also switches off the signalling pathway (Fig. 5B), showing
the essential role of the BceAB transporter in activating BceRS signalling in the absence of inducing signals. Typically, this TCS family confers resistance to antimicrobials targeting lipid
II cell wall metabolites such as nisin or bacitracin. Genetic activation of the pathway renders the BceSV124A mutant insensitive to nisin, which has a marked effect on the lag phase but not
on the growth rate of the WT strain, and increases resistance to gramicidin and, to a lesser extent, bacitracin (Fig. 5C and Supplementary Fig. 5). Interestingly, the BceSV124A mutant is
also more susceptible to antimicrobial peptides (colistin and polymyxin D) compared to the WT strain, while equally susceptible as the WT to vancomycin (Fig. 5C and Supplementary Fig. 5).
These results show that BceRS activation confers protection against structurally unrelated drugs targeting lipid II intermediates at the cost of increased susceptibility to antimicrobial
peptides. We then tested the phenotypes associated with loss-of-function of BceRS. Inactivation of the pathway in the BceSV124A background by additional BceRD55A or ∆_bceAB_ mutations leads
to nisin hyper-susceptibility while restoring WT level of colistin susceptibility (Fig. 5D). Nisin hyper-susceptibility is also observed for the single BceRD55A mutant (Fig. 5D), a phenotype
not linked to down expression of BceRS regulated genes (Fig. 5B). These results show that the BceRS system is constitutively active in the absence of drugs and that the basal activity is
necessary and sufficient to counteract the effects of sub-inhibitory concentrations of nisin. To test the current model of nisin resistance based on drug efflux and cleavage, we inactivated
the BceCDE transporter, the BceO substrate-binding protein, and the BceP extracellular protease (Fig. 5A). Deletion of ∆_bceD_ and ∆_bceO_ in the WT or BceSV124A backgrounds has no impact on
the nisin phenotypes of the respective parental strains (Fig. 5E), excluding a major function in drug export or binding. In contrast, the ∆_bceP_ mutant is slightly more susceptible to
nisin compared to the WT parental strain (Fig. 5E). However, deletion of _bceP_ in the BceSV124A background was always associated with secondary mutations inactivating the whole signalling
pathway in five independent mutants (Supplementary Data 2). Altogether, these results show that individual genes do not provide drug resistance and suggest that the BceP extracellular S41
protease56 has a buffering role when the pathway is activated, rather than directly cleaving drugs through an atypical mechanism, as previously suggested57. To test if BceRS regulates an
adaptive response rather than a resistance mechanism per se, we pre-incubated the WT strain with nisin for four hours. Prior exposure to the drug decreases the lag phase in a dose-dependent
manner upon subsequent exposure to higher nisin concentrations (Fig. 5F). For instance, adaptation with 8 µg/ml nisin, a WT sub-inhibitory concentration, confers a BceSV124A-like resistance
against a subsequent 64 µg/ml nisin challenge (Fig. 5F). More generally, prior adaptation with a given nisin concentration increases by a 4-fold factor the inhibiting concentration. These
results show that the BceRS response is adaptative and suggests that the BceRS system actively monitors and adjusts surface-exposed lipid II metabolites, rather than directly detoxifying
drugs or drug-lipid II complexes. DISCUSSION Our systematic analysis highlights the benefits of the HK+ gain-of-function approach to characterise TCS signalling, both for mapping regulatory
networks and for characterising individual systems. This study was made possible by the conserved mechanism of HK phosphatase activity originally proposed31,32, which allowed a single
residue to be targeted to activate the corresponding signalling pathway. By systematically testing all HisKA and HisKA_3 systems in a bacterium, we show the broad potential of this approach
to reveal specialised, connected, and global regulatory systems covering the functional diversity that has evolved from a simple two-component architecture. Targeting the HK phosphatase
catalytic residue has the advantage of leaving a quasi-native system. The gain-of-function is solely dependent on the HK mutation, with no change to the RR and preservation of the
physiological feedback loops. A second major advantage is that it bypasses the need for environmental signals, which are often unknown or confounding when having a wide effect on bacterial
physiology. In this respect, the SaeRS system is a remarkable example. Previous studies demonstrated SaeR regulation of _pbsP_ and _bvaP_ during vaginal colonisation, among transcriptomic
perturbations affecting nearly 40% of the genome49. However, the regulon remained elusive due to a lack of activation in vitro49. The HK+ approach resolves the signalling pathway by
revealing a specialised and CovR-connected, pathway. Comparison with the well-characterised _Staphylococcus aureus_ homologous system58 highlights the evolutionary divergence between
regulatory circuits, particularly for those regulating host-pathogen interactions, which need to be studied in each species. Originality is a mechanism linking the SaeRS and CovRS systems.
Complex regulatory wiring can be selected to mount co-ordinated responses, primary trough transcriptional cascades (a TCS regulating transcription of a second TCS) or connectors (usually a
TCS-regulated transmembrane protein modulating the activity of a second TCS)59. The C-peptide of adhesins can act as connectors when the transmembrane end remaining after cleavage of the
LPxTG motif by sortase A interacts with a histidine kinase53. The mechanism differs in GBS in which the PbsP adhesin domain acts as an extracellular signalling molecule to activate CovR
signalling, independently of cell wall anchoring. We hypothesise that the lysin-rich and positively charged PbsP adhesin interacts with CovS, with the co-regulatory proteins Abx1 and
Stk154,60, or with the negatively charged membrane, recalling the activation of the homologous CovRS system in _Streptococcus pyogenes_ by cationic peptides61. To complete the regulatory
circuit, CovR has previously been shown to repress _pbsP_ in a strain-specific manner45,50,51. The intertwining of SaeR and CovR signalling through PbsP constitutes an adaptive mechanism for
balancing adhesion and invasion and could contribute to the phenotypic and pathogenicity variabilities observed within the species. HK+ mutations resolve TCS regulatory networks but reveal
discrepancies in the activation of signalling pathways. While primary sequence analysis of TCSs did not uncover specific motifs correlating with high, intermediate, or low pathway
activation, two underlying factors may dampen the effect of HK+ mutations. First, HK kinase activity can be inhibited by interacting proteins, such as the small LiaF protein inhibiting
LiaS62,63 and the Pst/PhoU proteins inhibiting PhoRS64,65. Genes encoding co-regulatory proteins are often themselves regulated by the TCS, creating feedback loops that lock HKs in
kinase-deficient conformation and thus obliterate the effect of HK+ mutations. However, the presence of auto-inhibitory proteins is not a sufficient condition for preventing activation, as
demonstrated by VicK, which is inhibited by YycH/YycI66,67 but still activated by an HK+ mutation. Second, intermediary activation of the RelRS and CiaRH pathways suggests buffering
mechanisms for TCSs regulating multiple independent loci and integrated cellular response. However, detailed analysis is required to decipher phosphorylation dynamics in each
phosphatase-deficient HK+ and correlate in vivo RR phosphorylation with regulatory network activation, considering variable factors like the source of RR phosphorylation (kinase activity of
the HK+ variant, cross-talk by other HK, small metabolites) and specific spontaneous dephosphorylation rates of the labile aspartate residues27,68,69. The systematic approach validates the
conservation of the dephosphorylation mechanism. It also uncovers an unanticipated activation of the BceRS system with a degenerate QMKV motif. Recent structural insights from _Bacillus
subtilis_ complexes into the membrane environment support a highly dynamic model of interactions between the BceAB transporter and the BceS kinase- and phosphatase-competent
conformations70,71. Our results with the HK+ BceS indeed suggest that BceAB is necessary to stabilise the kinase-competent conformation of BceS. Alternatively, BceAB could also provide the
catalytic residue on the models suggested for the auxiliary phosphatases RapH and Spo0E31,72. At the phenotypic level, our results point towards a need-based mechanism of target protection,
as recently suggested for the Bce-like system73,74,75, and not towards a drug cleavage-exclusion mechanism as initially suggested76,77. The target protection mechanism relies on the binding
of lipid II intermediates on a binding pocket of BceAB70. However, it is still unclear how the system releases lipid II when it is complexed with drugs. Our results suggest an alternative
scenario in which BceAB constantly monitor free lipid II intermediate to minimise target exposure78,79. This alternative is supported by the steady-state activity of the BceRS pathway in the
absence of drugs and is compatible with a need-based mechanism. Further studies should test the entire BceRS pathway without relying on a lipid II-drug detoxification mechanism but rather
on a mechanism that maintains the steady-state level of free lipid II in the presence of drugs. It is also interesting to note that the activation of the BceRS-BceAB system impacts the
retention or secretion of the ß-h/c toxin, potentially as a result of the interaction between the polyene backbone of the toxin and the membranes46,47,48, suggesting functional links between
cell envelope homoeostasis, drug resistance, and virulence. To conclude, genetic activation by HK+ is a powerful approach to characterise positive regulation by TCS. It circumvents the
major drawback of studying systems that are usually non-activated in standard conditions. Previous studies on individual TCSs have demonstrated the potential of the approach, but it has
unfortunately not been widely adopted to date. Our systematic analysis based on the conserved mechanism of phosphatase activity provides a blueprint to decipher signalling, response
dynamics, evolution of gene regulation, and regulatory networks. The HK+ approach is recommended for the study of TCS in any species, either as a complement or as a first choice alongside a
classical deletion mutant. METHODS STRAIN, BACTERIAL GENETICS, AND GENOME SEQUENCING The BM110 strain is a clinical isolate representative of the hypervirulent CC-17 clonal complex
responsible for most neonatal meningitis80. The 2.2 Mb annotated genome is available under the NCBI RefSeq reference NZ_LT714196. The standard growth condition is in Todd-Hewitt medium
supplemented with 1% yeast extract and 50 mM Hepes pH 7.4 (THY) incubated in static condition at 37 °C. Oligonucleotides and construction of vectors for site-directed mutagenesis and
deletion are detailed in Supplementary Datas 5 and 6, respectively. Splicing-by-overlap PCR with high-fidelity polymerase (Thermo Scientific Phusion Plus) were done with complementary
primers containing the desired mutations. The final PCR products contain mutations (SNP or deletion) flanked on either side by 500 bp of sequence homologous to the targeted loci. Cloning is
done by Gibson assembly in pG1, a thermosensitive shuttle vector similar to the pG + host5 vector81. Constructs were introduced and maintained in _E. coli_ XL1-blue (Stratagene) with
erythromycin selection (150 µg/ml) at 37 °C. Inserts were validated by Sanger sequencing (Eurofins Genomics). Mutant construction in GBS was performed through a three-step process involving
episomal replication, chromosomal integration, and vector loss. Initially, pG vectors were introduced into GBS via electroporation, and transformants were selected on THY agar supplemented
with 5 µg/ml erythromycin at 30 °C, the permissive temperature for episomal replication. After 24–36 h of growth, two single transformant colonies were isolated on THY agar containing
erythromycin and incubated another 24–36 h at 37 °C, the non-permissive temperature for vector replication, thereby promoting chromosomal integration of the vector at the targeted locus
through homologous recombination. Subsequently, rare colonies that had integrated the vector into the chromosome were isolated on THY agar with erythromycin at 37 °C for 16 h. The isolated
integrants were inoculated into 10 ml of THY medium without antibiotic, incubated at 30 °C to activate the rolling circle origin of replication, and subjected to serial subculture twice
daily. By day 3, cultures were diluted (typically 10−5), spread onto THY and Columbia agar supplemented with 10% horse blood (BioMerieux), and incubated at 37 °C. Isolated colonies (_n_ =
24–48) were picked into 150 µl of THY in 96-well plates, incubated for 4–6 h at 37 °C, and replica-plated using a 96-pin replicator (Boekel Scientific) onto THY agar plates with and without
erythromycin. After 16 h at 37 °C, erythromycin-susceptible colonies that had lost the vector were identified and confirmed by discriminatory PCR (MyTaq HS - Bioline) using specific
oligonucleotides with the expected mutation at their 3’ extremity (Supplementary Data 5) to distinguish mutant from wild-type genotypes. Mutants were then isolated on THY at 37 °C, single
colonies inoculated in THY incubated at 37 °C for 16 h, centrifuged and resuspended in 20% glycerol for long-term storage at − 80 °C. Genomic DNA of at least two independent mutants for each
construction were purified from 1 ml of culture following manufacturer instruction for Gram-positive bacteria (DNeasy Blood and Tissue – Qiagen) and sequenced (Illumina sequencing at Core
facility or Eurofins Genomics). High-quality reads in Fastq were mapped against the BM110 genomes and analysed with Geneious Prime (2019.2.3 – Biomatters Ltd) using default parameters
(mapping: up to 5 iterations, minimum mapping quality Phred score 30, medium/low sensitivity; SNP calling: minimum variant frequency 0.25, maximum variant _P_-value 10−6, minimum strand-bias
_P_-value 10−5, with visual inspection for coverage and validation of SNPs). Results of genome sequencing for all mutants used in this study (55–419 x coverage, mean 181 x) are summarised
in Supplementary Data 2. RNA SEQUENCING RNA purification, sequencing and analysis were conducted essentially as described for the characterisation of the virulence regulator CovR45. The 14
HK+ mutants were split into two series of 8 strains (7 mutants and one WT strain), and RNA was purified using three independent replicates that were grown on different days. Overnight
cultures were used to inoculate THY (1/50), and 10 ml of culture were harvested in the exponential growth phase (OD600 = 0.5) after incubation at 37 °C. Bacterial pellets are washed with
cold PBS containing RNA stabilisation reagents (RNAprotect, Qiagen) before flash freezing and storage at − 80 °C. Total RNA are extracted after cell wall mechanical lysis with 0.1 µm beads
(Precellys Evolution, Bertin Technologies) in RNApro reagent (MP Biomedicals), and purified by chloroform extraction and ethanol precipitation. Samples were treated to remove residual DNA
(TURBO DNase, Ambion) before fluorescent-based quantification (Qubit RNA HS, Invitrogen) and quality validation (Agilent Bioanalyzer 2100). Depletion of rRNA (FastSelect Bacterial, Qiagen),
library construction and sequencing were done following manufacturer instructions (TruSeq Stranded mRNA, NextSeq 500, Illumina). Single-end strand-specific 75 bp reads were cleaned (cutadapt
v2.10) and mapped on the BM110 genome (Bowtie v2.5.1, with default parameters). Gene counts (featureCounts, v2.0.0, parameters: -t gene -g locus_tag -s 1) were analysed with R (v4.0.5) and
the Bioconductor package DESeq2 (v1.30.1)82. Normalisation, dispersion, and statistical tests for differential expression were performed with independent filtering. For each comparison, raw
_p_-values were adjusted using Benjamini and Hochberg multiple tests83 and adjusted _p_-values lower than 0.05 were considered significant. Raw sequencing reads and statistical analysis are
publicly available (GEO accession number GSE261394). In addition to HK+ RNA-sequencing, we have included an independent CovST282A transcriptome that was done simultaneously with the CovRD53A
transcriptome45, the latter being already reported altogether with CovR ChIP-seq experiment (GEO accession number GSE158049). Gene networks are represented with the open-source software
Cytoscape (v3.9.1)84. RT-QPCR AND PROMOTER ACTIVITY For validation, independent RNA purifications from biological triplicates were done using the same protocol, except that the cultures were
grown on the same day and only 1 ml was harvested. Reverse transcription and quantitative PCR (iScript Reverse Transcription and SsoAdvanced Universal SYBR Green, BioRad) were done using
specific primers (Supplementary Data 5). Fold changes are calculated for each target relative to the WT strain whose RNA was purified in parallel. For promoter activities, promoters were
amplified and cloned in the pTCV-lac vector containing a ß-galactosidase reporter (Supplementary Datas 5 and 6) and introduced in GBS. Reporter activity was quantified in microplate format
by colourimetric assay with ONPG as substrate and permeabilized overnight cultures54. Reaction kinetics at 28 °C were followed by OD at 420 nm every 5 min (Tecan Infinite). Linear slopes
(OD/min) were used to infer enzymatic activities and were normalised for the initial cell density (OD 600 nm) of each replicate. GROWTH CURVES AND ANTIBIOTIC SUSCEPTIBILITIES Growth curves
are done in a volume of 150 µl of THY inoculated with diluted overnight cultures (1/500) in 96-wells microplates and incubated at 37 °C with automatic recording of OD 600 nm every 10 minutes
and 1 min agitation by cycle (TECAN Infinite). Doubling times are determined by fitting non-linear regression with a Malthusian growth model (GraphPad Prism 10) in the exponential phase
(_R_2 > 0.99) for each replicate. Fitness is calculated by dividing the mean doubling time of the WT by the doubling time of the mutant. For antibiotic susceptibilities, concentrated
drugs (10 x) were added to an aliquot of the starting cultures and serial two-fold dilutions were done in the starting culture without drugs before incubation in the microplate reader.
Minimal Inhibitory Concentration (MIC) is done following EUCAST guidelines in Muller-Hinton Fastidious culture media (MH-F, Becton Dickinson) media using custom AST Sensititre 96 wells
plates (ThermoScientific) and 18 h of incubation at 37 °C. SS-HAEMOLYTIC ACTIVITY Columbia agar supplemented with 5% horse blood and Granada medium (BioMerieux) were used to visualise
ß-haemolytic activity and pigmentation, respectively. Serial ten-fold dilutions of cultures were spotted onto media plates and then incubated under anaerobic conditions (AnaeroGen, Oxoid) at
37 °C. To highlight the halo of lysis around colonies, images are converted to greyscale and uniformly processed (Photoshop, Adobe) to adjust contrast and brightness. Haemolytic titres were
determined by a semi-quantitative method54. Serial 2-fold dilution of cultures initially adjusted to 109 CFU/ml in PBS were added (V/V) to 1% defibrinated horse blood (Oxoid) in PBS
supplemented with 0.2% glucose. After 1 hour of incubation at 37 °C, cells were gently pelleted, and haemoglobin in supernatants was quantified by optical absorbance at 420 nm. The
haemolytic activity of each strain was defined as the minimum dilution that lysed at least 50% of red blood cells. Haemolytic titres are the ratio between the haemolytic activity of each
replicate against the haemolytic activity of the WT strain. Haemolytic titres are then normalised against the WT strain (normalised WT titre = 1). RR PHOSPHORYLATION LEVEL Genes encoding RR
were amplified and cloned by Gibson assembly (Supplementary Data 5 and 6) in a custom-made pEX-CterFLAG vector containing a synthetic cassette with a translational initiation site, a
flexible Gly-Ala linker, a 3xFLAG epitope, and a transcriptional terminator. Cassettes with genes of interest cloned in frame with the linker were excised with restriction enzymes and cloned
into the anhydro-tetracycline (aTc) inducible expression vector pTCV_PtetO45. Expression vectors were introduced in the corresponding HK+ mutants by electroporation with kanamycin
selection. Total protein extracts were prepared from 45 ml of cultures in exponential phase in the presence of 100 ng/ml aTc (Sigma) by mechanical lysis of bacterial pellet (Precellys
Evolution) resuspend in cold TBS buffer with EDTA-free protease inhibitors (cOmplete, Roche). Following clearance by centrifugation, 15 µg of proteins were loaded in 12.5% Phos-Tag SDS
polyacrylamide gels (SuperSep Phos-Tag, Wako Pure Chemical Industries Ltd) in loading dye buffer without EDTA and without sample heating to avoid dephosphorylation of the labile aspartate85.
Electrophoresis (2 h, 100 V, 30 mA) in Tris-glycine buffer was performed on an ice bath. Semi-dry transfer on nitrocellulose membrane (15 min, 15 V, Mini-Protean, BioRad) was followed by
blocking (TBS buffer with 0.05% Tween20 and 5% BSA), and hybridisation with rabbit polyclonal anti-FLAG antibodies (1:1500, Millipore F7425) and finally with secondary antibodies coupled to
infra-red dyes (1:15000, Li-Cor 926-32211 IRDye 800CW). After final washing in TBS buffer without Tween20, fluorescent signals were acquired (Odyssey Imager, Li-Cor). The ratio of
phosphorylated and non-phosphorylated proteins was analysed with ImageJ from three independent protein extracts. REPORTING SUMMARY Further information on research design is available in the
Nature Portfolio Reporting Summary linked to this article. DATA AVAILABILITY The RNA-seq data generated in this study have been deposited in the Gene Expression Omnibus database under
accession code GSE261394 [https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE261394]. Additional RNA-seq and ChIP-seq data already reported are available under accession code GSE261394
[https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE158049]. Source data are provided in this paper. REFERENCES * Stock, A. M., Robinson, V. L. & Goudreau, P. N. Two-component signal
transduction. _Annu. Rev. Biochem._ 69, 183–215 (2000). Article CAS PubMed Google Scholar * Groisman, E. A. Feedback control of two-component regulatory systems. _Annu. Rev. Microbiol._
70, 103–124 (2016). Article CAS PubMed PubMed Central Google Scholar * Groisman, E. A., Duprey, A. & Choi, J. How the PhoP/PhoQ system controls virulence and Mg(2+) homeostasis:
Lessons in signal transduction, pathogenesis, physiology, and evolution. _Microbiol. Mol. Biol. Rev._ 85, e0017620 (2021). Article PubMed Google Scholar * Francis, V. I. & Porter, S.
L. Multikinase networks: Two-component signaling networks integrating multiple stimuli. _Annu. Rev. Microbiol._ 73, 199–223 (2019). Article CAS PubMed Google Scholar * Sastry, A. V. et
al. The _Escherichia coli_ transcriptome mostly consists of independently regulated modules. _Nat. Commun._ 10, 5536 (2019). Article ADS CAS PubMed PubMed Central Google Scholar * Xie,
Y. et al. An atlas of bacterial two-component systems reveals function and plasticity in signal transduction. _Cell Rep._ 41, 111502 (2022). Article CAS PubMed Google Scholar * Capra,
E. J. & Laub, M. T. Evolution of two-component signal transduction systems. _Annu. Rev. Microbiol._ 66, 325–347 (2012). Article CAS PubMed PubMed Central Google Scholar * Salazar,
M. E. & Laub, M. T. Temporal and evolutionary dynamics of two-component signaling pathways. _Curr. Opin. Microbiol._ 24, 7–14 (2015). Article CAS PubMed PubMed Central Google Scholar
* Perez, J. C. & Groisman, E. A. Evolution of transcriptional regulatory circuits in bacteria. _Cell_ 138, 233–244 (2009). Article CAS PubMed PubMed Central Google Scholar *
Rapun-Araiz, B., Haag, A. F., Solano, C. & Lasa, I. The impact of two-component sensorial network in staphylococcal speciation. _Curr. Opin. Microbiol._ 55, 40–47 (2020). Article CAS
PubMed PubMed Central Google Scholar * Perez, J. C. & Groisman, E. A. Transcription factor function and promoter architecture govern the evolution of bacterial regulons. _Proc. Natl.
Acad. Sci. USA_ 106, 4319–4324 (2009). Article ADS CAS PubMed PubMed Central Google Scholar * Choi, J. & Groisman, E. A. Horizontally acquired regulatory gene activates ancestral
regulatory system to promote _Salmonella_ virulence. _Nucleic Acids Res._ 48, 10832–10847 (2020). Article CAS PubMed PubMed Central Google Scholar * Ghose, D. A., Przydzial, K. E.,
Mahoney, E. M., Keating, A. E. & Laub, M. T. Marginal specificity in protein interactions constrains evolution of a paralogous family. _Proc. Natl. Acad. Sci. USA_ 120, e2221163120
(2023). Article CAS PubMed PubMed Central Google Scholar * Rapun-Araiz, B. et al. Systematic reconstruction of the complete two-component sensorial network in _Staphylococcus aureus_.
_mSystems_ 5, e00511–e00520 (2020). Article CAS PubMed PubMed Central Google Scholar * Villanueva, M. et al. Sensory deprivation in _Staphylococcus aureus_. _Nat. Commun._ 9, 523
(2018). Article ADS PubMed PubMed Central Google Scholar * Galagan, J. E. et al. The _Mycobacterium tuberculosis_ regulatory network and hypoxia. _Nature_ 499, 178–183 (2013). Article
ADS CAS PubMed PubMed Central Google Scholar * Minch, K. J. et al. The DNA-binding network of _Mycobacterium tuberculosis_. _Nat. Commun._ 6, 5829 (2015). Article ADS CAS PubMed
Google Scholar * Trouillon, J. et al. Determination of the two-component systems regulatory network reveals core and accessory regulations across _Pseudomonas aeruginosa_ lineages. _Nucleic
Acids Res._ 49, 11476–11490 (2021). Article CAS PubMed PubMed Central Google Scholar * Gao, R. & Stock, A. M. Biological insights from structures of two-component proteins. _Annu.
Rev. Microbiol._ 63, 133–154 (2009). Article CAS PubMed PubMed Central Google Scholar * Jacob-Dubuisson, F., Mechaly, A., Betton, J. M. & Antoine, R. Structural insights into the
signalling mechanisms of two-component systems. _Nat. Rev. Microbiol._ 16, 585–593 (2018). Article CAS PubMed Google Scholar * Buschiazzo, A. & Trajtenberg, F. Two-component sensing
and regulation: How do histidine kinases talk with response regulators at the molecular level? _Annu. Rev. Microbiol._ 73, 507–528 (2019). Article CAS PubMed Google Scholar * Pioszak, A.
A. & Ninfa, A. J. Genetic and biochemical analysis of phosphatase activity of _Escherichia coli_ NRII (NtrB) and its regulation by the PII signal transduction protein. _J. Bacteriol._
185, 1299–1315 (2003). Article CAS PubMed PubMed Central Google Scholar * Dutta, R., Yoshida, T. & Inouye, M. The critical role of the conserved Thr247 residue in the functioning of
the osmosensor EnvZ, a histidine Kinase/Phosphatase, in _Escherichia coli_. _J. Biol. Chem._ 275, 38645–38653 (2000). Article CAS PubMed Google Scholar * Hsing, W., Russo, F. D., Bernd,
K. K. & Silhavy, T. J. Mutations that alter the kinase and phosphatase activities of the two-component sensor EnvZ. _J. Bacteriol._ 180, 4538–4546 (1998). Article CAS PubMed PubMed
Central Google Scholar * Kenney, L. J. How important is the phosphatase activity of sensor kinases? _Curr. Opin. Microbiol._ 13, 168–176 (2010). Article CAS PubMed PubMed Central
Google Scholar * Laub, M. T. & Goulian, M. Specificity in two-component signal transduction pathways. _Annu. Rev. Genet._ 41, 121–145 (2007). Article CAS PubMed Google Scholar *
Gao, R. & Stock, A. M. Quantitative kinetic analyses of shutting off a two-component system. _MBio_ 8, e00412–e00417 (2017). Article CAS PubMed PubMed Central Google Scholar *
Trajtenberg, F. et al. Regulation of signaling directionality revealed by 3D snapshots of a kinase:regulator complex in action. _Elife_ 5, e21422 (2016). Article PubMed PubMed Central
Google Scholar * Casino, P., Rubio, V. & Marina, A. Structural insight into partner specificity and phosphoryl transfer in two-component signal transduction. _Cell_ 139, 325–336 (2009).
Article CAS PubMed Google Scholar * Skerker, J. M. et al. Rewiring the specificity of two-component signal transduction systems. _Cell_ 133, 1043–1054 (2008). Article CAS PubMed
PubMed Central Google Scholar * Huynh, T. N. & Stewart, V. Negative control in two-component signal transduction by transmitter phosphatase activity. _Mol. Microbiol._ 82, 275–286
(2011). Article CAS PubMed PubMed Central Google Scholar * Huynh, T. N., Noriega, C. E. & Stewart, V. Conserved mechanism for sensor phosphatase control of two-component signaling
revealed in the nitrate sensor NarX. _Proc. Natl. Acad. Sci. USA_ 107, 21140–21145 (2010). Article ADS CAS PubMed PubMed Central Google Scholar * Willett, J. W. & Kirby, J. R.
Genetic and biochemical dissection of a HisKA domain identifies residues required exclusively for kinase and phosphatase activities. _PLoS Genet._ 8, e1003084 (2012). Article CAS PubMed
PubMed Central Google Scholar * Mikkelsen, H., Hui, K., Barraud, N. & Filloux, A. The pathogenicity island encoded PvrSR/RcsCB regulatory network controls biofilm formation and
dispersal in _Pseudomonas aeruginosa_ PA14. _Mol. Microbiol._ 89, 450–463 (2013). Article CAS PubMed PubMed Central Google Scholar * Horstmann, N. et al. Phosphatase activity of the
control of virulence sensor kinase CovS is critical for the pathogenesis of group A _Streptococcus_. _PLoS Pathog._ 14, e1007354 (2018). Article PubMed PubMed Central Google Scholar *
Wayne, K. J., Li, S., Kazmierczak, K. M., Tsui, H. C. & Winkler, M. E. Involvement of WalK (VicK) phosphatase activity in setting WalR (VicR) response regulator phosphorylation level and
limiting cross-talk in _Streptococcus pneumoniae_ D39 cells. _Mol. Microbiol._ 86, 645–660 (2012). Article CAS PubMed PubMed Central Google Scholar * Sharkey, L. K. R. et al. The
two-component system WalKR provides an essential link between cell wall homeostasis and DNA replication in _Staphylococcus aureus_. _mBio_ 14, e0226223 (2023). Article PubMed Google
Scholar * Armistead, B., Oler, E., Adams Waldorf, K. & Rajagopal, L. The double life of group B _Streptococcus_: asymptomatic colonizer and potent pathogen. _J. Mol. Biol._ 431,
2914–2931 (2019). Article CAS PubMed PubMed Central Google Scholar * Vornhagen, J., Adams Waldorf, K. M. & Rajagopal, L. Perinatal group B streptococcal infections: virulence
factors, immunity, and preventionstrategies. _Trends Microbiol._ 25, 919–931 (2017). Article CAS PubMed PubMed Central Google Scholar * Thomas, L. & Cook, L. Two-component signal
transduction systems in the human pathogen _Streptococcus agalactiae_. _Infect. Immun._ 88, e00931–19 (2020). Article PubMed PubMed Central Google Scholar * Dobihal, G. S., Brunet, Y.
R., Flores-Kim, J. & Rudner, D. Z. Homeostatic control of cell wall hydrolysis by the WalRK two-component signaling pathway in _Bacillus subtilis_. _Elife_ 8, e52088 (2019). Article CAS
PubMed PubMed Central Google Scholar * Monk, I. R., Howden, B. P., Seemann, T. & Stinear, T. P. Spontaneous secondary mutations confound analysis of the essential two-component
system WalKR in _Staphylococcus aureus_. _Nat. Commun._ 8, 14403 (2017). Article ADS CAS PubMed PubMed Central Google Scholar * Winkler, M. E. & Hoch, J. A. Essentiality, bypass,
and targeting of the YycFG (VicRK) two-component regulatory system in gram-positive bacteria. _J. Bacteriol._ 190, 2645–2648 (2008). Article CAS PubMed PubMed Central Google Scholar *
Joubert, L. et al. Visualization of the role of host heme on the virulence of the heme auxotroph _Streptococcus agalactiae_. _Sci. Rep._ 7, 40435 (2017). Article ADS CAS PubMed PubMed
Central Google Scholar * Mazzuoli, M. V. et al. The CovR regulatory network drives the evolution of Group B _Streptococcus_ virulence. _PLoS Genet._ 17, e1009761 (2021). Article CAS
PubMed PubMed Central Google Scholar * Whidbey, C. et al. A hemolytic pigment of Group B _Streptococcus_ allows bacterial penetration of human placenta. _J. Exp. Med._ 210, 1265–1281
(2013). Article CAS PubMed PubMed Central Google Scholar * Armistead, B. et al. Lipid analogs reveal features critical for hemolysis and diminish granadaene mediated Group B
_Streptococcus_ infection. _Nat. Commun._ 11, 1502 (2020). Article ADS CAS PubMed PubMed Central Google Scholar * Armistead, B. et al. Hemolytic membrane vesicles ofgroup B
_Streptococcus_ promote infection. _J. Infect. Dis._ 223, 1488–1496 (2021). Article CAS PubMed Google Scholar * Cook, L. C., Hu, H., Maienschein-Cline, M. & Federle, M. J. A vaginal
tract signal detected by the GBS SaeRS system elicits transcriptomic changes and enhances murine colonization. _Infect. Immun._ 86, e00762–17 (2018). Article PubMed PubMed Central Google
Scholar * Buscetta, M. et al. PbsP, a cell wall-anchored protein that binds plasminogen to promote hematogenous dissemination of group B _Streptococcus_. _Mol. Microbiol._ 101, 27–41
(2016). Article CAS PubMed Google Scholar * Lentini, G. et al. The plasminogen binding protein PbsP is required for brain invasion by hypervirulent CC17 Group B streptococci. _Sci. Rep._
8, 14322 (2018). Article ADS PubMed PubMed Central Google Scholar * Thomas, L. S. & Cook, L. C. A novel conserved protein in _Streptococcus agalactiae_, BvaP, is important for
vaginal Colonization and Biofilm Formation. _mSphere_ 7, e0042122 (2022). Article PubMed Google Scholar * Hall, J. W. et al. An intramembrane sensory circuit monitors sortase A-mediated
processing of streptococcal adhesins. _Sci. Signal._ 12, eaas9941 (2019). Article PubMed PubMed Central Google Scholar * Firon, A. et al. The Abi-domain protein Abx1 interacts with the
CovS histidine kinase to control virulence gene expression in group B _Streptococcus_. _PLoS Pathog._ 9, e1003179 (2013). Article CAS PubMed PubMed Central Google Scholar * Mascher, T.
Bacterial (intramembrane-sensing) histidine kinases: signal transfer rather than stimulus perception. _Trends Microbiol._ 22, 559–565 (2014). Article CAS PubMed Google Scholar *
Sommerfield, A. G. & Darwin, A. J. Bacterial carboxyl-terminal processing proteases play critical roles in the cell envelope and beyond. _J. Bacteriol._ 204, e0062821 (2022). Article
PubMed Google Scholar * Khosa, S. et al. Structural basis of lantibiotic recognition by the nisin resistance protein from _Streptococcus agalactiae_. _Sci. Rep._ 6, 18679 (2016). Article
ADS CAS PubMed PubMed Central Google Scholar * Jenul, C. & Horswill, A. R. Regulation of _staphylococcus aureus_ virulence. _Microbiol. Spectr._ 7,
https://doi.org/10.1128/microbiolspec.GPP3-0031-2018 (2019). * Mitrophanov, A. Y. & Groisman, E. A. Signal integration in bacterial two-component regulatory systems. _Genes Dev._ 22,
2601–2611 (2008). Article CAS PubMed PubMed Central Google Scholar * Lin, W. J. et al. Threonine phosphorylation prevents promoter DNA binding of the Group B _Streptococcus_ response
regulator CovR. _Mol. Microbiol._ 71, 1477–1495 (2009). Article CAS PubMed PubMed Central Google Scholar * Gryllos, I. et al. Induction of group A _Streptococcus_ virulence by a human
antimicrobial peptide. _Proc. Natl. Acad. Sci. USA_ 105, 16755–16760 (2008). Article ADS CAS PubMed PubMed Central Google Scholar * Jani, S. et al. Low phosphatase activity of LiaS and
strong LiaR-DNA affinity explain the unusual LiaS to LiaR in vivo stoichiometry. _BMC Microbiol._ 20, 104 (2020). Article CAS PubMed PubMed Central Google Scholar * Sanson, M. A. et
al. The LiaFSR transcriptome reveals an interconnected regulatory network in group A _streptococcus_. _Infect. Immun._ 89, e0021521 (2021). Article PubMed Google Scholar * Lamarche, M.
G., Wanner, B. L., Crepin, S. & Harel, J. The phosphate regulon and bacterial virulence: a regulatory network connecting phosphate homeostasis and pathogenesis. _FEMS Microbiol. Rev._
32, 461–473 (2008). Article CAS PubMed Google Scholar * Baek, S. & Lee, E. J. PhoU: a multifaceted regulator in microbial signaling and homeostasis. _Curr. Opin. Microbiol._ 77,
102401 (2024). Article CAS PubMed Google Scholar * Szurmant, H., Bu, L., Brooks III, C. L. & Hoch, J. A. An essential sensor histidine kinase controlled by transmembrane helix
interactions with its auxiliary proteins. _Proc. Natl. Acad. Sci. USA_ 105, 5891–5896 (2008). Article ADS CAS PubMed PubMed Central Google Scholar * Szurmant, H., Mohan, M. A., Imus,
P. M. & Hoch, J. A. YycH and YycI interact to regulate the essential YycFG two-component system in _Bacillus subtilis_. _J. Bacteriol._ 189, 3280–3289 (2007). Article CAS PubMed
PubMed Central Google Scholar * Gao, R. & Stock, A. M. Probing kinase and phosphatase activities of two-component systems in vivo with concentration-dependent phosphorylation
profiling. _Proc. Natl. Acad. Sci. USA_ 110, 672–677 (2013). Article ADS CAS PubMed Google Scholar * Butcher, R. J. & Tabor, J. J. Real-time detection of response regulator
phosphorylation dynamics in live bacteria. _Proc. Natl. Acad. Sci. USA_ 119, e2201204119 (2022). Article CAS PubMed PubMed Central Google Scholar * George, N. L., Schilmiller, A. L.
& Orlando, B. J. Conformational snapshots of the bacitracin sensing and resistance transporter BceAB. _Proc. Natl. Acad. Sci. USA_ 119, e2123268119 (2022). Article CAS PubMed PubMed
Central Google Scholar * George, N. L. & Orlando, B. J. Architecture of a complete Bce-type antimicrobial peptide resistance module. _Nat. Commun._ 14, 3896 (2023). Article ADS CAS
PubMed PubMed Central Google Scholar * Parashar, V., Mirouze, N., Dubnau, D. A. & Neiditch, M. B. Structural basis of response regulator dephosphorylation by Rap phosphatases. _PLoS
Biol._ 9, e1000589 (2011). Article CAS PubMed PubMed Central Google Scholar * Fritz, G. et al. A new way of sensing: Need-based activation of antibiotic resistance by a flux-sensing
mechanism. _mBio_ 6, e00975 (2015). Article CAS PubMed PubMed Central Google Scholar * Kobras, C. M. et al. BceAB-Type antibiotic resistance transporters appear to act by target
protection of cell wall synthesis. _Antimicrob. Agents Chemother._ 64, e02241–19 (2020). Article CAS PubMed PubMed Central Google Scholar * Koh, A., Gibbon, M. J., Van der Kamp, M. W.,
Pudney, C. R. & Gebhard, S. Conformation control of the histidine kinase BceS of _Bacillus subtilis_ by its cognate ABC-transporter facilitates need-based activation of antibiotic
resistance. _Mol. Microbiol._ 115, 157–174 (2021). Article CAS PubMed Google Scholar * Reiners, J. et al. The N-terminal region of Nisin is important for the BceAB-type ABC transporter
NsrFP from _streptococcus agalactiae_ COH1. _Front. Microbiol._ 8, 1643 (2017). Article PubMed PubMed Central Google Scholar * Yang, Y. et al. Role of two-component system response
regulator bceR in the antimicrobial resistance, virulence, biofilm formation, and stress response of group B _streptococcus_. _Front. Microbiol._ 10, 10 (2019). Article CAS PubMed PubMed
Central Google Scholar * Piepenbreier, H., Diehl, A. & Fritz, G. Minimal exposure of lipid II cycle intermediates triggers cell wall antibiotic resistance. _Nat. Commun._ 10, 2733
(2019). Article ADS PubMed PubMed Central Google Scholar * Kingston, A. W., Zhao, H., Cook, G. M. & Helmann, J. D. Accumulation of heptaprenyl diphosphate sensitizes _Bacillus
subtilis_ to bacitracin: implications for the mechanism of resistance mediated by the BceAB transporter. _Mol. Microbiol._ 93, 37–49 (2014). Article CAS PubMed PubMed Central Google
Scholar * Da Cunha, V. et al. _Streptococcus agalactiae_ clones infecting humans were selected and fixed through the extensive use of tetracycline. _Nat. Commun._ 5, 4544 (2014). Article
PubMed Google Scholar * Biswas, I., Gruss, A., Ehrlich, S. D. & Maguin, E. High-efficiency gene inactivation and replacement system for gram-positive bacteria. _J. Bacteriol._ 175,
3628–3635 (1993). Article CAS PubMed PubMed Central Google Scholar * Love, M. I., Huber, W. & Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with
DESeq2. _Genome Biol._ 15, 550 (2014). Article PubMed PubMed Central Google Scholar * Benjamini, Y. & Hochberg, Y. Controlling the false discovery rate: A practical and powerful
approach to multiple testing. _J. R. Stat. Soc. Ser. B_ 57, 289–300 (1995). Article MathSciNet Google Scholar * Shannon, P. et al. Cytoscape: a software environment for integrated models
of biomolecular interaction networks. _Genome Res._ 13, 2498–2504 (2003). Article CAS PubMed PubMed Central Google Scholar * Barbieri, C. M. & Stock, A. M. Universally applicable
methods for monitoring response regulator aspartate phosphorylation both in vitro and in vivo using Phos-tag-based reagents. _Anal. Biochem._ 376, 73–82 (2008). Article CAS PubMed PubMed
Central Google Scholar Download references ACKNOWLEDGEMENTS This study was supported by Agence Nationale de la Recherche (VirEvol - ANR-22-CE15-0024) to A.F., and the National Laboratory
of Excellence programme - Integrative Biology of Emerging Infectious Diseases (LabEx IBEID, ANR-10-LABX-62-IBEID) to P.T.C. C.C. and M.V.M. are recipients of National PhD grants from Ecole
Doctorale BioSpc (ED562) - Université Paris Cité. AUTHOR INFORMATION Author notes * These authors contributed equally: Cosme Claverie, Francesco Coppolino. AUTHORS AND AFFILIATIONS *
Institut Pasteur, Université Paris Cité, Department of Microbiology, Biology of Gram-Positive Pathogens, Paris, France Cosme Claverie, Francesco Coppolino, Maria-Vittoria Mazzuoli, Odile
Sismeiro, Patrick Trieu-Cuot & Arnaud Firon * University of Messina, Department of Human Pathology, Messina, Italy Francesco Coppolino, Giuseppe Valerio De Gaetano & Concetta
Beninati * Assistance Publique-Hôpitaux de Paris, Hôpital Cochin, Department of Bacteriology, French National Reference Center for Streptococci, Paris, France Cécile Guyonnet & Asmaa
Tazi * Université Paris Cité, Institut Cochin, Institut National de la Santé et de la Recherche Médicale U1016, Centre National de la Recherche Scientifique UMR8104, Team Bacteria and
Perinatality, Paris, France Cécile Guyonnet & Asmaa Tazi * Fédération Hospitalo-Universitaire Fighting Prematurity, Paris, France Cécile Guyonnet & Asmaa Tazi * Institut Pasteur,
Université Paris Cité, Bioinformatics and Biostatistics Hub, Paris, France Elise Jacquemet & Rachel Legendre * Scylla Biotech Srl, Messina, Italy Giuseppe Teti Authors * Cosme Claverie
View author publications You can also search for this author inPubMed Google Scholar * Francesco Coppolino View author publications You can also search for this author inPubMed Google
Scholar * Maria-Vittoria Mazzuoli View author publications You can also search for this author inPubMed Google Scholar * Cécile Guyonnet View author publications You can also search for this
author inPubMed Google Scholar * Elise Jacquemet View author publications You can also search for this author inPubMed Google Scholar * Rachel Legendre View author publications You can also
search for this author inPubMed Google Scholar * Odile Sismeiro View author publications You can also search for this author inPubMed Google Scholar * Giuseppe Valerio De Gaetano View
author publications You can also search for this author inPubMed Google Scholar * Giuseppe Teti View author publications You can also search for this author inPubMed Google Scholar * Patrick
Trieu-Cuot View author publications You can also search for this author inPubMed Google Scholar * Asmaa Tazi View author publications You can also search for this author inPubMed Google
Scholar * Concetta Beninati View author publications You can also search for this author inPubMed Google Scholar * Arnaud Firon View author publications You can also search for this author
inPubMed Google Scholar CONTRIBUTIONS C.C., F.C., M.V.M., C.G., O.S., and G.V.D.G. performed experiments and analysed data. E.J. and R.L. analysed RNA-seq data. G.T., P.T.C., A.T., C.B., and
A.F. designed and supervised experiments. A.F. conceived the study and wrote the manuscript with input from all the authors. CORRESPONDING AUTHOR Correspondence to Arnaud Firon. ETHICS
DECLARATIONS COMPETING INTERESTS GT is an employee and CB is the founder and owner of Scylla Biotech Srl. The company did not provide funding and had no role in the design, conduct, or
publication of the study. All other authors declare no competing interests. PEER REVIEW PEER REVIEW INFORMATION _Nature Communications_ thanks Taeok Bae and the other anonymous reviewers for
their contribution to the peer review of this work. A peer review file is available. ADDITIONAL INFORMATION PUBLISHER’S NOTE Springer Nature remains neutral with regard to jurisdictional
claims in published maps and institutional affiliations. SUPPLEMENTARY INFORMATION SUPPLEMENTARY INFORMATION PEER REVIEW FILE DESCRIPTION OF ADDITIONAL SUPPLEMENTARY FILES SUPPLEMENTARY DATA
1 SUPPLEMENTARY DATA 2 SUPPLEMENTARY DATA 3 SUPPLEMENTARY DATA 4 SUPPLEMENTARY DATA 5 SUPPLEMENTARY DATA 6 REPORTING SUMMARY SOURCE DATA SOURCE DATA RIGHTS AND PERMISSIONS OPEN ACCESS This
article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as
you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party
material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s
Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder.
To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. Reprints and permissions ABOUT THIS ARTICLE CITE THIS ARTICLE Claverie, C., Coppolino, F., Mazzuoli, MV.
_et al._ Constitutive activation of two-component systems reveals regulatory network interactions in _Streptococcus agalactiae_. _Nat Commun_ 15, 9175 (2024).
https://doi.org/10.1038/s41467-024-53439-3 Download citation * Received: 04 April 2024 * Accepted: 07 October 2024 * Published: 24 October 2024 * DOI:
https://doi.org/10.1038/s41467-024-53439-3 SHARE THIS ARTICLE Anyone you share the following link with will be able to read this content: Get shareable link Sorry, a shareable link is not
currently available for this article. Copy to clipboard Provided by the Springer Nature SharedIt content-sharing initiative