Effect of reward on electrophysiological signatures of grid cell population activity in human spatial navigation
- Select a language for the TTS:
- UK English Female
- UK English Male
- US English Female
- US English Male
- Australian Female
- Australian Male
- Language selected: (auto detect) - EN
Play all audios:

The regular equilateral triangular periodic firing pattern of grid cells in the entorhinal cortex is considered a regular metric for the spatial world, and the grid-like representation
correlates with hexadirectional modulation of theta (4–8 Hz) power in the entorhinal cortex relative to the moving direction. However, researchers have not clearly determined whether grid
cells provide only simple spatial measures in human behavior-related navigation strategies or include other factors such as goal rewards to encode information in multiple patterns. By
analysing the hexadirectional modulation of EEG signals in the theta band in the entorhinal cortex of patients with epilepsy performing spatial target navigation tasks, we found that this
modulation presents a grid pattern that carries target-related reward information. This grid-like representation is influenced by explicit goals and is related to the local characteristics
of the environment. This study provides evidence that human grid cell population activity is influenced by reward information at the level of neural oscillations.
Grid cells have a periodic equilateral triangle firing pattern, which is considered the basis for their metric measurement in spatial navigation1. Grid cells with different scales of firing
fields can provide accurate positioning information through scale combination2. This internal metric helps organisms achieve path integration and vector-based navigation behaviors such as
homing in the absence of external information3. Since it is the universal metric system for spatial navigation, the grid pattern should be rigid and immutable. However, based on the in-depth
study of grid cells, the grid pattern is not strictly unchanged but can be squeezed, stretched, deformed or even discretely broken under the effects of various types of environmental
spatial information4,5,6 and can also be merged globally after multiple subenvironmental spaces are connected7. This result is a step forward in the understanding of grid cells, which
provide flexible metric systems that adapt to the spatial environment and assist organisms in completing path integration in complex spatial environments. Recently, some breakthrough studies
on the information types coded by grid cells have been published. In 2016, Tim Behrens et al. found that grid cells also present a typical hexadirectional modulation mode when encoding
abstract bird space with nonspatial information8. In the same year, Neil Burgess and his team also discovered the hexadirectional modulation of grid cells for the encoding of imaginary
space9. Afterward, the discussion and research on nonphysical spatial information (such as conceptual space, social space, etc.) encoded by grid cells were initiated10,11. Thus, the grid
cells in the entorhinal cortex appear to encode a broader “cognitive map” in a more general sense, with their distinctive equilateral triangular periodicity patterns12. Since grid cells can
encode both spatial and nonspatial information, will they be affected by nonspatial information when encoding the physical environment? Boccara et al. and Butler et al. independently
conducted experiments on rodents to address this question in 201913,14. The results of both studies show that the grid pattern is indeed affected by nonspatial information such as goal
reward, resulting in local deformation and structural adjustment. For the exploration of nonspatial information encoded by grid cells, particularly high-level decision-making information
such as navigation goals, the more important research object is humans, but experimental research on humans is very scarce. Limited by experimental conditions, research on changes in the
human grid pattern is often limited to noninvasive functional magnetic resonance imaging (fMRI) measurements, and the signal from the entorhinal cortex in the limbic system where grid cells
are located is difficult to record. Here, we performed a VR desktop navigation experiment on patients with medically intractable epilepsy and recorded EEG data from deep electrodes in the
entor
hinal cortex to investigate the nonspatial effect of grid cell population activity. Based on recent research showing that theta oscillations in the human entorhinal cortex carry
hexadirectional modulation information of grid cells15,16, we also used the same method to study the changes in grid cell population activity by measuring theta power. The hexadirectional
modulation of theta power, which reflects the regular activity of grid cells, only appears in the experimental stage without an effect of a specific goal object. However, when a clear goal
object is present, the hexadirectional modulation of theta power disappears, indicating that the grid pattern is disturbed by the target reward. In addition, the disturbance is also affected
jointly by environmental boundaries, and in the central region lacking boundary anchor cues, it will depend more on the self-centered localization function of grid cells. Our study is the
first to analyse the effect of nonspatial information on a grid pattern in human spatial navigation by recording EEG signals from intracranial electrodes. This study provides mesoscopic
evidence for exploring the underlying coding patterns of the human entorhinal cortex and the coupling effect of spatial and nonspatial factors on the entorhinal cortex and advances the
research on the multidimensional generalized functional framework of grid cells.
Electroencephalograms were recorded from stereotactically implanted electrodes in patients with medically intractable epilepsy, and their seizure foci were located to guide their respective
treatment. The mean age of the 9 patients (3 females) was 27.1 years (SD = 8.2).
The Scientific Research Ethics Committee of Beijing Normal University approved all procedures performed in this study. Written informed consent was obtained from all patients. All the
experiments were performed in accordance with relevant guidelines and regulations.
The participants navigated freely in a circular virtual arena adapted by Doeller et al.17 using a laptop to perform a task of remembering the location of goal objects that included rewards.
The environment consists of a meadow plane (9500 virtual units in diameter) surrounded by a wall with a circular boundary. The navigation task paradigm was written using UnrealEngine 2 (Epic
Games). During the initial learning phase at the start of the experiment, patients were asked to remember the locations of eight different everyday objects. For about 10 min, participants
will traverse the real locations of the eight objects they will see in the experiment one by one, picking them up according to the objects presented in the scene. Patients then completed
different numbers of trials. Each trial consisted of cue, retrieval, feedback, and recoding phases (Fig. 1A). During the cue phase, the participant viewed one of the objects (for 2 s).
During the retrieval phase, they used the arrow keys (left, right, and forward) on the laptop keyboard to navigate to the location of the relevant object. The duration of this phase is a
self-determined step. When the participants reached what they thought was the correct position, they pressed the space bar to locate the object (Fig. 1B). Depending on the accuracy of the
response, the patient received feedback from one of five possible cartoon faces (lasting 1.5 s). The more red and sad the cartoon face was, the greater the error in behavior. The actual
reward received by the participants after completing the experiment will be given according to the behavior score, which means that the participants used the cartoon face as a cue to
determine how much reward they would receive. Then, the object appeared in the correct position, and the patient navigated to that position for further learning. Behavioral events and motion
data were written to log files with a temporal resolution of 10 ms. The patient was asked to complete more than 100 trials but could be instructed to pause or withdraw from the task at any
time.
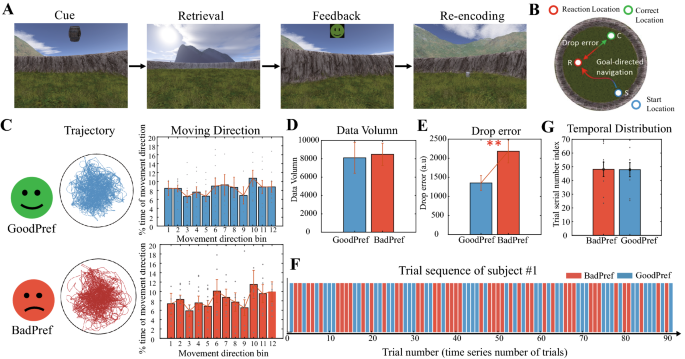
Paradigm and behavioral division with the effect of goals. (A) The four task stages of each trial in the memory task performed by the participants (adapted from Chen et al. (Current Biology,
2018)). (B) An aerial view of the circular arena. The drop error is defined as the relative distance between the subjective response location and the reallocation. (C) Two types of groups
are defined by drop error in behavior. The top row is GoodPerf, and the bottom row is BadPerf. The middle column is the trajectory of the participants, achieving extensive coverage in both
groups. The right-most column shows the movement direction of the participants, which is evenly distributed at all angles between the two groups (Rayleigh’s tests for non-uniformity, p >
0.05). (D) Comparison of experimental data between GoodPerf and BadPerf. A significant difference was not observed between the two groups (t test, p > 0.5). (E) Comparison of performance in
the trials corresponding to GoodPerf and BadPerf goal objects. The difference in drop error between the two groups was significant (t test, p