Common juniper, an overlooked conifer with high invasion potential in protected areas of Patagonia
- Select a language for the TTS:
- UK English Female
- UK English Male
- US English Female
- US English Male
- Australian Female
- Australian Male
- Language selected: (auto detect) - EN
Play all audios:

Although the benefits of early detection of incipient invasion in natural habitats are well recognized, so are the difficulties associated with it18,19,20. Detection of early invasion foci
is usually fortuitous16, and citizen collaboration is important in increasing the probability of registering these situations. Public engagement is being enhanced by collaborative projects,
led by professional scientists, that seek to compile information on potentially invasive species20,21. In particular, citizen science has emerged as a powerful tool for detecting and mapping
the distribution of recent invasive species and obtaining diverse bio-ecological information on them20,22,23. This knowledge can provide insights into the invasion stage, the mechanisms
behind the invasion, and the invader’s potential ecological impact, which can be context-dependent24,25.
Climate is recognized as the single most important factor determining the distribution of plant species at a large scale26,27. Thus, a frequently used approach to predict where a species
might invade is analysis of the climatic similarity between its native range and areas outside it28,29, even for plant species with no invasive history30. This approach has been used for
invasion risk assessment of non-native conifer species in areas of their introduced ranges throughout the Southern Hemisphere28,30, where they pose a significant threat to the diversity and
functioning of native ecosystems31 and even PAs32,33. In particular, climate matching can be a valuable tool for estimating suitable areas for potentially damaging non-native conifers with
incipient invasion. By cross-referencing information, it is possible to prioritize the search for and control of new invasion foci in, for example, PAs with high invasion risk.
The PAs of Andean Patagonia are no exception in terms of their high vulnerability to an increasing number of introduced plant species34,35. In this region, increasing anthropogenic pressure
on PAs acts as a catalyst for new invasions of introduced plant species whose invasive status, ecology, impact, and distribution are mostly unknown. This is exemplified by the conifer
Juniperus communis L. (native to temperate regions of the boreal hemisphere), which has been identified as a potential high-risk invader of climatically suitable areas in Africa30 and
Oceania28. At the southernmost tip of South America, Argentina, J. communis has recently been officially cataloged as an invasive species (Ministerio de Ambiente y Desarrollo Sostenible
2021). Despite this, no studies have addressed its invasion status, distribution, or potential expansion range, especially in the areas of the country where it may represent a risk to
biodiversity, such as the PAs. This species already has three validated records in PAs of Andean Patagonia, according to the Biodiversity Information System which provides biological
information on the species, and PAs of Argentina (www.sib.gob.ar). However, J. communis can be frequently seen in PAs of Andean Patagonia, which suggests that it is under-recorded, probably
because of its incipient invasion (i.e. the earliest stage of the invasion process). This assumption of an incipient invasion is supported by the lack of J. communis registers in key
reference literature describing the regional flora36, including literature focusing on introduced plant species in the main PAs of the region35,37. In addition, this species is increasingly
valued as the raw material for producing gin, an alcoholic beverage that is booming internationally. This encourages its cultivation in the area, which can increase the source of propagules
for invasions in nearby PAs. It can also be seen in gardens; however, its incidence as an ornamental plant, and therefore the importance of this type of use as a source of propagules, is as
yet unknown.
Here, we characterized the invasion status of the non-native conifer J. communis in PAs and interface areas of Andean Patagonia, Argentina, by mapping its distribution and describing both
the invasion and the environments this species inhabits. We registered the type of invaded habitats, species abundance, its spatial configuration pattern, the accompanying woody species, the
species’ reproductive potential (i.e. presence of reproductive plants and seedlings), its importance as an ornamental plant, and its occurrence in PAs and associated areas. We also modeled
the potential distribution of the species by comparing the climatic conditions in its introduced range in Patagonia with those of its native distribution range. We used different
methodological approaches to acquire data on the species in the region: a literature search, field sampling, and citizen collaboration. To our knowledge, this is the first work to provide
information on J. communis as an invader of a South American country. We address key descriptive aspects of the current J. communis invasion that provide clues to the ecological mechanisms
involved in its spread. Knowledge of the potential distribution of J. communis could be useful in determining the invasion risk the species presents for high conservation value ecosystems of
Patagonia.
We compiled 174 occurrences of J. communis in the region (58.6% from field sampling, 33.9% from citizen contributions, and 7.4% from the literature review); > 90% of these records were from
PAs (Fig. 1). We detected the presence of J. communis within eight PAs and close to another seven in the region (Table 1). Almost 100% of occurrences (sampled or reported) were associated
with disturbed environments, mostly represented by roadsides (gravel or paved) and trails (Fig. 2).
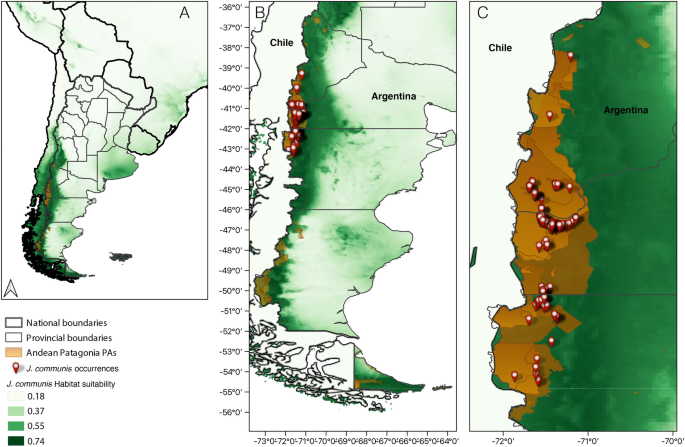
Juniperus communis occurrences (red symbols) in PAs and urban-natural interface areas (orange area) of Andean Patagonia, Argentina panels (B) and (C). In the three panels the potential
distribution model for J. communis generated by Maxent is shown, panel (A) shows a general view of Argentina and neighboring countries, panel (B) shows the entire Patagonia and panel (C)
shows a zoom to the sampled area. Habitat suitability is represented on a green scale, with darker colors representing higher suitability. In (C), the largest lighter orange area represents
the World Biosphere Reserve, which overlaps most of the other PAs. The map was created using QGIS version 3.28.2-Firenze (www.qgis.org).
A. Steppe invasion, B. Forest, walking trail invasion, and C. High-abundance shrubland invasion. D. Roadside invasion. E. Co-occurrence with a native woody species (Diostea juncea). F.
Co-occurrence with a non-native woody species (Pinus contorta). G. Mature (purple) and immature (green) fruits on the same individual. H. Juniperus communis’ hedge. I. Ornamental tree. The
photographs in this research work were captured by the authors at diverse locations spanning the study area.
Field sampling data indicated that J. communis was found most frequently in forests (48%), followed by shrublands (26%), with the lowest representation in steppe environments (5%; Fig. 2).
We found an equal occurrence of the species in natural and urban habitats, with only a small percentage of ornamental use (Figs. 2, 3). Regarding the spatial distribution pattern, J.
communis was frequently found as isolated individuals (62%), followed by thickets (21%) and, to a lesser extent, both patterns in the same site (5%; Figs. 2, 3). The species was found mostly
at low abundance (2–10 individuals in 45% of the occurrences), followed by a single individual (24%), medium density (11–100 individuals in 16% of the occurrences), and high density (> 100
individuals in 11% of the occurrences; Figs. 2, 3). Fruited individuals and seedlings were observed in ca. 70%, and 50% of the registers, respectively (Fig. 3), which could represent an
underestimation of seedling presence since the understory of some sites was difficult to explore due to dense vegetation. In addition, we registered 24 main woody species accompanying J.
communis, half of which were native (Fig. 4). The most frequently found native species were Austrocedrus chilensis (Cupressaceae), Maytenus boaria (Celastraceae), Nothofagus dombeyi
(Nothofagaceae), and Lomatia hirsuta (Proteaceae). Among the non-native species the most frequently found were Pinus contorta (Pinaceae), Rosa rubiginosa (Rosaceae), and Cytisus scoparius
(Fabaceae) (Figs. 1, 4).
Descriptive variables related to J. communis and the environments it inhabits. The graph depicts the proportion of registers for different sub-categories according to habitat (steppe,
shrubland, forest, other; n = 87), environment (natural, rural, urban; n = 114) if the species was used as ornamental (n = 161), species abundance (single, low, medium, and high; n = 73),
the spatial configuration pattern of the individuals (thicket, isolated, both; n = 88), and the presence of fruits (n = 103) and seedlings (n = 72).
Frequency of the principal woody species that accompanied J. communis occurrences (n = 38). The X-axis represents the proportion of sites where each species was observed, with a maximum
value of 0.15 indicating that these species were found in 15% of the sampled sites. Bar colors indicate species origin: native (green) or introduced (orange). The donut figure shows the
percentage of occurrences for native and introduced species.
The literature review afforded 18 records of J. communis cited as a naturally established species in the Andean Patagonian region (Neuquén, Río Negro, and Chubut provinces), with the oldest
record dating back to 2002 (Table 2). Most of the records corresponded to PAs (82%), including four national parks. In most of the studies the inclusion of J. communis was not intentional,
rather it appeared when describing vegetation, or when listing introduced species (Table 2). Only three studies considered the species as the focus of their research, although none of these
recognized it as invasive (Table 2).
The Andean Patagonian region showed a highly climatically suitable land area for J. communis occurrence (Figs. 1, 5), with the area of the best fitting model covering all major PAs in the
region (Fig. 1). The area of greatest suitability occupies the region near the Andes, from central to southern Argentina, becoming longitudinally wider towards the north of Andean Patagonia
and extending eastward into the southern part of Río Negro and northern Chubut.
Comparison of environmental variables between the PAs of Patagonia and the native range of J. communis: mean temperature of coldest and warmest quarters (top panel), and mean precipitation
of coldest and warmest quarters (bottom panel). The Y-axis represents the estimated probability density. In all cases a P-value 100), the spatial configuration of the individuals (thicket,
isolated, or both), the main woody species, whether the species was occurring naturally or not (e.g. ornamental), and if there were individuals with fruits and seedlings (