Resilience to periodic disturbances and the long-term genetic stability in acropora coral
- Select a language for the TTS:
- UK English Female
- UK English Male
- US English Female
- US English Male
- Australian Female
- Australian Male
- Language selected: (auto detect) - EN
Play all audios:

ABSTRACT Climate change is restructuring natural ecosystems. The direct impacts of these events on biodiversity and community structure are widely documented, but the impacts on the genetic
variation of populations remains largely unknown. We monitored populations of _Acropora_ coral on a remote coral reef system in northwest Australia for two decades and through multiple
cycles of impact and recovery. We combined these demographic data with a temporal genetic dataset of a common broadcast spawning corymbose _Acropora_ to explore the spatial and temporal
patterns of connectivity underlying recovery. Our data show that broad-scale dispersal and post-recruitment survival drive recovery from recurrent disturbances, including mass bleaching and
mortality. Consequently, genetic diversity and associated patterns of connectivity are maintained through time in the broader metapopulation. The results highlight an inherent resilience in
these globally threatened species of coral and showcase their ability to cope with multiple disturbances, given enough time to recover is permitted. SIMILAR CONTENT BEING VIEWED BY OTHERS
DANGEROUS DEMOGRAPHICS IN POST-BLEACH CORALS REVEAL BOOM-BUST VERSUS PROTRACTED DECLINES Article Open access 22 September 2021 DISPARATE GENETIC DIVERGENCE PATTERNS IN THREE CORALS ACROSS A
PAN-PACIFIC ENVIRONMENTAL GRADIENT HIGHLIGHT SPECIES-SPECIFIC ADAPTATION Article Open access 07 July 2023 GLOBAL WARMING DECREASES CONNECTIVITY AMONG CORAL POPULATIONS Article 30 December
2021 INTRODUCTION The capacity of natural populations to recover from acute disturbances and maintain genetic variation is a key aspect of their resilience1,2,3,4. The process of recovery is
primarily influenced by rates of survival, growth and immigration5,6. When mortality is high and immigration is low, recovery depends on the few individuals surviving locally, and declines
in genetic diversity are expected7. In contrast, when immigration is high and impacts vary among sub-populations, areas least affected can serve as a source of recruits to aid in the
recovery more widely. In these cases, genetic diversity remains stable8. On tropical coral reefs, acute heat stress events and associated bleaching have become routine disturbances and are
now the primary cause for coral mortality globally9,10. Monitoring patterns of recovery and associated changes in coral cover have informed how climate change is impacting coral
reefs11,12,13; however, information on how fluctuating coral populations influence genetic variation are also required. The isolated oceanic atolls of northwest Australia provide a unique
opportunity to study coral populations through cycles of impact and recovery. The Scott system of three reefs is hundreds of kilometres from the mainland and neighbouring reefs, and is
demographically isolated in space and time14,15,16,17,18. In recent decades, recurrent disturbances to the Scott reefs have caused dramatic shifts in coral population sizes and
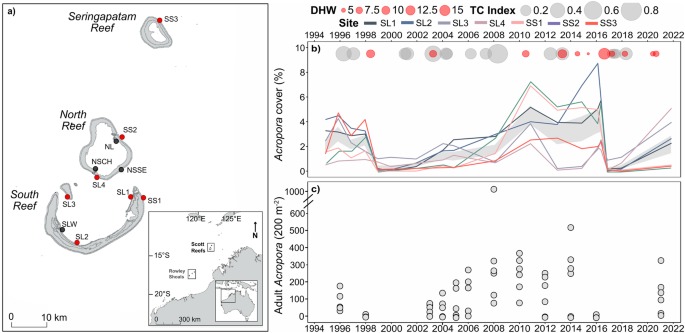
distribution11,19,20,21. From 1994 to 2021, moderate heat stress and tropical cyclones were frequent disturbances, reducing coral cover at one or more of our long-term monitoring (LTM) sites
during 9 of the 27 years (Fig. 1). The most severe disturbances were mass bleaching events in 1998, and again in 2016, causing relative reductions in mean coral cover of ~75% following both
events11. Cyclone Lua in 2012 also decreased mean cover by 8% across the reef system22, with other smaller disturbances (e.g. bleaching in 2010) causing less severe and more localised
impacts. The reefs had largely recovered 12 years after the 1998 mass bleaching, but with local variation in recovery depending on subsequent exposure to disturbances and the resilience of
different coral groups within communities, leading to long-term shifts in community composition11. RESULTS AND DISCUSSION Through the regime of recent disturbances to the Scott system of
reefs, the impacts of even the most severe event varied spatially, usually with a different group of sites worst affected by bleaching or by cyclones (Fig. 1b). Consequently, when _Acropora_
cover decreased to very low levels (<1%) at some sites following disturbance, it was often higher (1–7%) at other sites across the reef system (Fig. 1b). This spatial variation in cover
of _Acropora_ reflected the changes in abundance of adult colonies (>20 cm) within the long-term monitoring sites (Fig. 1c). The loss of adult colonies caused a comparative reduction in
reproductive output and larval supply to recruitment tiles across the reef system (R2 = 0.857; red points, Fig. 2a); however, this stock-recruitment relationship was much weaker at the scale
of the individual sites (200 m2), with the abundance of adult colonies being a poor predictor of larval supply to settlement tiles locally (R2 = 0.438; purple points, Fig. 2a). Subsequent
recovery at each site was then driven by local conditions, with the supply of _Acropora_ larvae to settlement tiles predicting (R2 = 0.706) the number of new _Acropora_ recruits (<2 cm)
on natural substrata six to 12 months later (Fig. 2b). Survival of these recruits and the resulting juveniles (6–15 cm) was also high (Fig. 2c), with the number of recruits on substrata
influencing the number of juvenile colonies within each site 12–24 months later (Fig. 2d). Demographic changes in _Acropora_ assemblages provided indirect evidence of recovery at the Scott
reefs being seeded by the supply of larvae from less affected sites. This mechanism of recovery was supported by population genetics of a common species of corymbose _Acropora_. Low-coverage
whole genome re-sequencing of _Acropora sp_. (formerly _Acropora tenuis_23) revealed high gene flow and connectivity across the Scott system of reefs, with no clustering of samples by site
using a genotype likelihood approach based on 8,329,442 variant sites (Fig. 3a). This pattern of high gene flow was confirmed using more stringent depth filters (29,462 single nucleotide
polymorphisms (SNPs); Fig. S1). Mean _F_ST was −0.0002 (Fig. S1) and most pairwise estimates between sites were negative and few were significant (Fig. 3b). The exception was the sample site
from Seringapatam reef (SS3), which showed low (FST < 0.004) but significant levels of genetic differentiation with all other sites from North and South Scott (Fig. 3b, Supplementary
Data 1). Together, the whole genome re-sequencing data pointed to broadscale dispersal and high gene flow across the reef system. Our temporal genetic dataset of microsatellite genotypes
(2004, 2009, 2014, 2015, 2021) revealed a consistent pattern of connectivity underlying recovery (Fig. 3c). Within each year, mean estimates of genetic differentiation were negative, and
most pairwise values were low and non-significant (Fig. S2, Supplementary Data 2). The genetic patchiness that did arise within each timepoint was ephemeral and changed from year to year
(Fig. S3). As a result, there were no significant differences in allele frequencies among sites nested within years (θST = −0.008, _P_ = 0.752). Differences among years under the AMOVA
framework accounted for only 1.6% of the total variation (θRT = 0.0192, _P_ = 0.003) and we did not observe any clustering of samples by year in multi-dimensional space (Supplementary Data
3, Fig. 3c). This pattern of chaotic genetic patchiness is a common attribute of broadcast spawning marine species, where the complex interplay between biological and physical processes
often drives unstable patterns of spatial genetic structure24,25. In turn, one generation of patches does not predict the next (Fig. S4). These levels of genetic differentiation among sites
and years were outside the bounds of normality when we randomized site-time assignments and recalculated genetic differentiation (Fig. S5), suggesting that they more likely reflect
stochastic biological processes than technical artefacts. Despite recurrent disturbances and dramatic demographic changes in _Acropora_ assemblages, the genetic diversity of the broader
metapopulation at the Scott reefs has remained stable. We did not observe any changes in allelic richness or heterozygosity among our sampling timepoints (Fig. 4a, b; Supplementary Data 4).
While site-level estimates seemed to vary slightly among years, genetic diversity in the broader population remained stable across our study period. Although we did not have samples from the
Scott reefs that predated the 1999 mass bleaching event, we found that levels of genetic diversity were similar to samples collected from the neighbouring Rowley Shoals, an offshore oceanic
atoll system 400 km to the south of the Scott reefs that has avoided widespread bleaching and mortality over similar timescales11. Indeed, microsatellite markers (Fig. 4a, b, Supplementary
Data 4) and whole genome re-sequencing techniques (Fig. 4c, d) indicated that genetic diversity in the two systems are similar, despite strong genetic drift (genome-wide FST = 0.078, Fig.
S6) and contrasting disturbance histories. Thus, despite severe declines in coral cover, our data suggest that there have been enough survivors at Scott system of reefs to facilitate
recovery and maintain moderate levels of genetic diversity in the metapopulation. The persistence of coral populations depends largely on their connectivity and larval supply, and their
capacity to absorb and adapt to emerging pressures. Our results suggest that a high dispersal capacity and post-recruitment survival in _Acropora_ are key mechanisms of resilience to climate
change. This is in contrast to species with a lower capacity to disperse across a group of reefs, such as some brooding corals or those reliant on vegetative propagation26. However, our
data also highlight that recovery depends on spatial variation in impacts and relatively high abundances of survivors at several locations across the reef system, in addition to considerable
time between severe disturbances. Under these conditions, population abundances and genetic diversity of _Acropora_ corals may be maintained, but with an increasing severity and frequency
of disturbances through global heating, longer term degradation is likely, particularly at these isolated coral reefs. This study underscores the importance of long-term monitoring to
understand the impacts of climate change on coral reef ecosystem health. More importantly, it highlights an inherent resilience in broadcast spawning coral populations on remote reefs, and
offers a glimmer of hope to coral reef ecosystems more broadly against a backdrop of rapid environmental change. METHODS DEMOGRAPHIC CHANGES IN _ACROPORA_ In 1994, permanent transects (250
metres) were established at 21 reef slope sites replicated across seven locations and three reefs27. At each location (Fig. 1a), three sites were separated by approximately 300 metres and
consisted of 250 metres of permanent transects marked at 10-m intervals. Surveys were conducted annually between 1994 and 1999, and then in 2003, 2004, 2005, 2008, 2010, 2012, 2014, 2016,
2017, and 2021. In 2016, additional surveys were conducted in January, April and October 2016, before, during and after the mass bleaching, and from 2016 only the first site at each location
was surveyed to allow for expansion into additional habitats (not included here). During each survey, a tape was laid along the permanent transect and images of the benthic community
captured from a distance between 30 and 50 cm from the substrata. Images were analysed using point sampling technique and benthic groups identified to the lowest taxonomic resolution
achievable by each observer28. These data were then divided among benthic groups according to taxa (e.g., family, genus) and growth form (e.g. encrusting, foliose, massive, branching).
LARVAL SUPPLY AND RECRUITMENT Larval supply of the most diverse and abundant coral genus, the _Acropora_, was quantified using terracotta settlement tiles along the permanent transects at 18
sites at six of the reef slope locations, during nine years between 1996 to 201321 (1996–1999, 2002, 2003, 2008, 2010, 2011). At each of the sites, groups of six settlement tiles (10 cm ×
10 cm) were deployed at distances of 30–100 cm apart, spaced at 60 m along the permanent transect. Settlement tiles (_n_ = 108 tiles total) were deployed and collected one month either side
of the primary mass coral spawning29. Size-frequency distributions of _Acropora_ colonies (excluding staghorn and hispidose growth forms) were quantified along the permanent transects (200
m) at the first site at each of six reef slope locations. Colony sizes (longest linear dimension) were recorded to the nearest centimetre along ten 20 m transects spaced at 10 m intervals,
within a width of 25 cm for colonies <10 cm (50 m2) and 1 m for colonies ≥10 cm (200 m2). Density at each site was adjusted to 200 m2. Growth and survival of colonies was quantified in a
species of corymbose _Acropora_ (nominally _Acropora spicifera_) at two of the permanent sites at four reef slope locations over four years (2006–2010). Locations were chosen to be
representative of the primary habitat for corymbose _Acropora_ across the reef system. Colonies (_n_ = 3692; of all sizes were tagged and resurveyed annually. Given issues identifying
juvenile coral to species, an assemblage of _Acropora_ species were tagged for colonies <10 cm in size. Size and growth were quantified from photographs taken directly above the colony,
or perpendicular to the maximum length when first tagged. Colonies were digitized and their size defined by the maximum diameter through the ellipse of best fit. MICROSATELLITE GENOTYPING
Tissue samples were collected from 982 colonies of a corymbose _Acropora (Acropora sp.)_ at five timepoints (2004, 2009, 2014, 2015, and 2021; Supplementary Data 5) spanning 17 years. At
each site, samples were taken from colonies separated by at least 1.5 m and not from loose fragments or colonies directly down-slope from another colony. Genomic DNA was extracted using a
Qiagen DNeasy kit or using a high-throughput membrane-based DNA extraction protocol30 and genotyped across at seven microsatellite loci16,31 in 10 µL multiplex PCR reactions using the Qiagen
multiplex PCR kit. Fragment analysis was carried out on a GE Healthcare MegaBACE 1000 capillary sequencer or an Applied Biosystems 3730 capillary sequencer. Electropherograms were
visualised and scored using the software GeneMarker v1.91 (SoftGenetics). To facilitate comparison of allele sizes among collections that were analysed on different platforms, 19 control
samples from earlier collections that exhibited a representative range of allele sizes were run alongside the new samples and allele sizes calibrated accordingly. To further minimise
genotyping errors, automated scorings of alleles in all microsatellite analyses were checked manually, and uncertainties were cleared by re-amplification and comparison. Of the seven loci
originally trialled, two loci scored in 2004 and 2009 (Amil2_006 and Amil5_028) could not be consistently scored and/or calibrated on the Applied Biosystems sequencer, and these loci were
removed. CRYPTIC LINEAGES AND CLONALITY Previous data from the Scott reefs have shown cryptic lineages are widespread in _Acropora_ and reflect divergence in reproductive timing, with the
main spawning period during austral autumn and a secondary smaller event in austral spring29,32. Sympatric colonies of _Acropora sp_. display strong genetic divergence, suggesting pronounced
reproductive boundaries and limited gene flow29,33,34,35. Before analysing patterns of genetic variation through space and time, an initial screening of all samples using microsatellite
data was conducted to remove the less abundant spring spawning lineage to focus our analyses on a single gene pool. To do this, we utilized data from 2009 that tracked a subset of colonies
(_n_ = 68) across multiple years with a consistent seasonal spawning phenotype29. These samples were used to assign samples from all timepoints to one of the two lineages using model-based
Bayesian clustering in Structure36 (Fig. S7). We ran _structure_ with the _no prior_ model at K = 2 based on the panel of five common microsatellites. Analyses were run ten times using the
admixture model with independent allele frequencies and with a burn-in period of 100,000, followed by 500,000 MCMC replications for each run. These model conditions were implemented because
the genetic differentiation between spawning groups is large and initial investigations showed that runs with correlated allele frequencies model overestimated K and individuals lacked
strong assignment29. After removing the spring spawning lineage and poor-quality samples that failed to amplify, we were left with 506 autumn spawning colonies (321 multi-locus genotypes)
with greater than 0.75 probability of assignment (mean assignment was 0.96 +/− 0.05 S.D.). Genotypic richness in the targeted species at the Scott reefs is high (>0.9615) with little
evidence of clonal propagation via vegetative fragmentation; however, replicate multi-locus genotypes were common in our temporal dataset when we restricted analyses to the five
microsatellite loci that could be scored across all years. The 2004 and 2009 datasets included two additional loci15 and showed that the replicate MLGs in the temporal dataset were indeed
unique individuals, and so we proceeded with a dataset of all 506 multi-locus genotypes for downstream analyses. We also clone corrected at the population level using _poppr_ and confirmed
patterns identified with the larger dataset held true when focussing on unique MLGs. GENETIC DIFFERENTIATION AND DIVERSITY Using our temporal dataset of microsatellite genotypes for the
autumn spawning lineage, we calculated pairwise estimates of _F_ST among sites with _stammp_37 and adjusted for multiple comparisons using a False Discovery Rate in R (_p.adjust_). We used
_prcomp_ (R Core Team, 2021) to carry out principal components analyses and used Discriminant Analyses of Principal Components (DAPC) with _adegenet_38 to estimate group assignment
probabilities under a Bayesian framework. To explore hierarchical patterns of genetic structure, we carried out an analysis of molecular variance (AMOVA) with sample sites nested within
timepoints. To measure levels of genetic diversity, we calculated allelic richness and expected heterozygosity for each site and timepoint comparison using _hierfstat_39. We then used a
Bayesian generalised linear mixed effects model with a gaussian distribution to test for differences in genetic diversity among years. Sampling site was included as a random effect to
account for the lack of spatial independence. The model was based on three chains with 3000 iterations, including 1000 iterations to warm-up and a thinning interval of 5. All Bayesian models
were performed in Stan (Stan Development Team, 2021) using the “brms” package40. We ran posterior predictive checks and visually inspected all models for violations for statistical
assumptions using the ‘dharma’ package (Hartig, 2022). Finally, to compare these estimates of genetic diversity with a pristine reef system, we genotyped conspecific colonies of _Acorpora
sp._ from the Rowley Shoals using the same panel of microsatellite markers (_n_ = 196 colonies) and whole genome re-sequencing techniques (_n_ = 10 colonies, below). WHOLE GENOME
RE-SEQUENCING We re-sequenced the genomes of samples (_n_ = 111 colonies) collected in 2021 to approximately 8X coverage (Fig. S8) using Covaris library preparation via sonication and
sequenced on a NovaSeq at Genomics WA. Raw sequence files were mapped with _bwa mem_ (Supplementary Note 1) to the _A. tenuis_ pseudo-chromosome-scale assembly (_aten.chr.fasta_)33. We used
_samtools_41 to sort and index and _picardtools_ (http://broadinstitute.github.io/picard) to mark and remove duplicate reads. _Samtools_41 was used to calculate sequencing depth of each
sample at each position across the genome (Supplementary Note 2, Fig. S10). We called single nucleotide polymorphisms (SNPs) using _mpileup_ in _bcftools_42 and filtered using _samtools_ and
based on a minor allele frequency of 0.05, a call rate of 0.95, and a mean read depth of 10x per sample (Supplementary Note 3). This produced a variant call file of 29,462 biallelic SNPs
that was used to estimate pairwise _F_ST among sites (minimum of 5 individuals per site) using _stamp_. We also explored genetic structure among sites using a genotype likelihood approach in
_angsd_43 and estimated genome-wide allele frequencies to carry out principal components analyses using _pcangsd_44 (Supplementary Note 4). We masked sites with a minQ < 20, minMapQ <
30, sites with less than 1/3 and greater than 2x the mean read depth and removed loci with missing data in more than half of the samples. We only retained variable sites (snp_pval <
1e−6) and those with a minor allele frequency greater than 0.05. This filtering criteria produced 8,329,442 variant sites used for clustering analysis. We also re-sequenced a subset of
genomes from the Rowley Shoals (_n_ = 10) using Nextera Flex Library Preparation Kits (Illumina) and sequenced on a NovaSeq. REPORTING SUMMARY Further information on research design is
available in the Nature Portfolio Reporting Summary linked to this article. DATA AVAILABILITY All of the data to support the findings of this study can be found on the Open Science Framework
https://doi.org/10.17605/OSF.IO/2VBHW (https://osf.io/2vbhw/)45. REFERENCES * Gunderson, L. H. Ecological resilience-in theory and application. _Annu. Rev. Ecol. Syst._ 31, 425–3 (2000).
Article Google Scholar * O’Neill, R. V. Recovery in complex ecosystems. _J. Aquat. Ecosyst. Stress Recover._ 6, 181–187 (1999). Article Google Scholar * Holling, C. S. Resilience and
stability of ecological systems. _Annu. Rev. Ecol. Syst._ 4, 1–23 (1973). Article Google Scholar * Nyström, M., Folke, C. & Moberg, F. Coral reef disturbance and resilience in a
human-dominated environment. _Trends Ecol. Evol._ 15, 413–417 (2000). Article PubMed Google Scholar * Davies, I. D., Cary, G. J., Landguth, E. L., Lindenmayer, D. B. & Banks, S. C.
Implications of recurrent disturbance for genetic diversity. _Ecol. Evol._ 6, 1181–1196 (2016). Article PubMed PubMed Central Google Scholar * Banks, S. C. et al. How does ecological
disturbance influence genetic diversity? _Trends Ecol. Evol._ 28, 670–679 (2013). Article PubMed Google Scholar * Beheregaray, L. B. et al. Genes record a prehistoric volcano eruption in
the Galápagos. _Science_ 302, 75 (2003). Article CAS PubMed Google Scholar * Spear, S. F., Crisafulli, C. M. & Storfer, A. Genetic structure among coastal tailed frog populations at
Mount St. Helens is moderated by post-disturbance management. _Ecol. Appl._ 22, 856–869 (2012). Article PubMed Google Scholar * Hughes, T. et al. Coral reefs in the Anthropocene. _Nature_
546, 82–90 (2017). Article CAS PubMed Google Scholar * Hughes, T. et al. Global warming and recurrent mass bleaching of corals. _Nature_ 543, 373–377 (2017). Article CAS PubMed
Google Scholar * Gilmour, J. P. et al. A tale of two reef systems: local conditions, disturbances, coral life histories, and the climate catastrophe. _Ecol. Appl_.
https://doi.org/10.1002/eap.2509 (2021). * Hoegh-guldberg, O. et al. Coral reefs under rapid climate change and ocean acidification. _Science_ 318, 1737–1742 (2007). Article CAS PubMed
Google Scholar * Pandolfi, J. M. et al. Global trajectories of the long-term decline of coral reef ecosystems. _Science_ 301, 955–958 (2003). Article CAS PubMed Google Scholar * Moore,
Ja. Y. et al. Unprecedented mass bleaching and loss of coral across 12° of latitude in Western Australia in 2010-11. _PLoS ONE_ 7, e51807 (2012). Article CAS PubMed PubMed Central Google
Scholar * Underwood, J. N. Genetic diversity and divergence among coastal and offshore reefs in a hard coral depend on geographic discontinuity and oceanic currents. _Evol. Appl._ 2,
222–233 (2009). Article PubMed PubMed Central Google Scholar * Underwood, J. N., Smith, L. D., van Oppen, M. J. H. & Gilmour, J. P. Ecologically relevant dispersal of corals on
isolated reefs: implications for managing resilience. _Ecol. Appl._ 19, 18–29 (2009). Article PubMed Google Scholar * Adam, A. A. S. et al. Diminishing potential for tropical reefs to
function as coral diversity strongholds under climate change conditions. _Divers. Distrib_. 27, 2245–2261 (2021). * Zhang, J. et al. Evolutionary responses of a reef-building coral to
climate change at the end of the last glacial maximum. _Mol. Biol. Evol._ 39, 1–18 (2022). Article Google Scholar * Smith, L. D., Gilmour, J. P. & Heyward, A. J. Resilience of coral
communities on an isolated system of reefs following catastrophic mass-bleaching. _Coral Reefs_ 27, 197–205 (2007). Article Google Scholar * Gilmour, J. P. et al. The state of Western
Australia’s coral reefs. _Coral Reefs_ 38, 651–667 (2019). Article Google Scholar * Gilmour, J. P., Smith, L. D., Heyward, A. J., Baird, A. H. & Pratchett, M. S. Recovery of an
isolated coral reef system following severe disturbance. _Science_ 340, 69–71 (2013). Article PubMed Google Scholar * Puotinen, M. et al. Towards modelling the future risk of cyclone wave
damage to the world’s coral reefs. _Glob. Chang. Biol._ 26, 4302–4315 (2020). Article PubMed Google Scholar * Bridge, T. C. L. et al. Original Article A tenuis relationship: traditional
taxonomy obscures systematics and biogeography of the ‘Acropora tenuis’ (Scleractinia: Acroporidae) species complex. _Zool. J. Linn. Soc_. 27, 1–24 (2023). * Johnson, M. & Black, R.
Chaotic genetic patchiness in an intertidal limpet, Siphonaria sp. _Mar. Biol._ 164, 157–164 (1982). Article Google Scholar * Johnson, M. & Black, R. Pattern beneath the chaos: the
effect of recruitment on genetic patchiness in an intertidal limpet. _Evolution_ 38, 1371–1383 (1984). Article PubMed Google Scholar * Thomas, L. et al. Contrasting patterns of genetic
connectivity in brooding and spawning corals across a remote atoll system in northwest Australia. _Coral Reefs_ 39, 55–60 (2020). Article Google Scholar * Heyward, A. et al.
_Characterisation of Scott Reef Lagoon Biota – Fish and Macrobenthos_ (1999). * Jonker, M., Johns, K. & Osborne, K. _Surveys of Benthic Reef Communities Using Underwater Digital
Photography and Counts of Juvenile Corals_. _Long-term Monitoring of the Great Barrier Reef_, Vol. 10 (2008). * Gilmour, J. P., Underwood, J. N., Howells, E. J., Gates, E. & Heyward, A.
J. Biannual spawning and temporal reproductive isolation inacropora corals. _PLoS ONE_ 11, 1–14 (2016). Article Google Scholar * Ivanova, N. V., Dewaard, J. R. & Hebert, P. D. N. An
inexpensive, automation-friendly protocol for recovering high-quality DNA. _Mol. Ecol. Notes_ 6, 998–1002 (2006). Article CAS Google Scholar * van Oppen, M. J. H., Underwood, J. N.,
Muirhead, A. & Peplow, L. Ten microsatellite loci for the reef-building coral Acropora millepora (Cnidaria, Scleractinia) from the Great Barrier Reef. _Aust. Mol. Ecol._ 7, 436–438
(2007). Google Scholar * Gilmour, J. P., Smith, L. D. & Brinkman, R. M. Biannual spawning, rapid larval development and evidence of self-seeding for scleractinian corals at an isolated
system of reefs. _Mar. Biol._ 156, 1297–1309 (2009). Article Google Scholar * Thomas, L. et al. Spatially varying selection between habitats drives physiological shifts and local
adaptation in a broadcast spawning coral on a remote atoll in Western Australia. _Sci. Adv._ 8, abl9185 (2022). Article Google Scholar * Rosser, N. L. et al. Phylogenomics provides new
insight into evolutionary relationships and genealogical discordance in the reef-building coral genus _Acropora_. _Proc. R. Soc. B Biol. Sci._ 284, 20162182 (2017). Article Google Scholar
* Rosser, N. L., Edyvane, K., Malina, A. C., Underwood, J. N. & Johnson, M. S. Geography and spawning season drive genetic divergence among populations of the hard coral Acropora tenuis
from Indonesia and Western Australia. _Coral Reefs_ 39, 989–999 (2020). Article Google Scholar * Pritchard, J. K., Stephens, M. & Donnelly, P. Inference of population structure using
multilocus genotype data. _Genetics_ 155, 945–959 (2000). Article CAS PubMed PubMed Central Google Scholar * Pembleton, L. W., Cogan, N. O. I. & Forster, J. W. StAMPP: an R package
for calculation of genetic differentiation and structure of mixed-ploidy level populations. _Mol. Ecol. Resour._ 13, 946–952 (2013). Article CAS PubMed Google Scholar * Jombart, T.
Adegenet: a R package for the multivariate analysis of genetic markers. _Bioinformatics_ 24, 1403–1405 (2008). Article CAS PubMed Google Scholar * Goudet, J. HIERFSTAT, a package for R
to compute and test hierarchical F-statistics. _Mol. Ecol. Notes_ 5, 184–186 (2005). Article Google Scholar * Bürkner, P. C. Bayesian Item Response Modeling in R with brms and Stan. _J.
Stat. Softw_. 100, 1–54 (2021). * Li, H. et al. The sequence alignment/map format and SAMtools. _Bioinformatics_ 25, 2078–2079 (2009). Article PubMed PubMed Central Google Scholar * Li,
H. A statistical framework for SNP calling, mutation discovery, association mapping and population genetical parameter estimation from sequencing data. _Bioinformatics_ 27, 2987–2993 (2011).
Article CAS PubMed PubMed Central Google Scholar * Korneliussen, T. S., Albrechtsen, A. & Nielsen, R. ANGSD: analysis of next generation sequencing data. _BMC Bioinforma._ 15, 1–13
(2014). Article Google Scholar * Meisner, J. & Albrechtsen, A. Inferring population structure and admixture proportions in low-depth NGS data. _Genetics_ 210, 719–731 (2018). Article
PubMed PubMed Central Google Scholar * Thomas, L. Resilience to periodic disturbances and the long-term genetic stability in Acropora coral. _Open Sci. Framew_.
https://doi.org/10.17605/OSF.IO/2VBHW (2024). Download references ACKNOWLEDGEMENTS This research was funded by the Australian Institute of Marine Science and Woodside Energy Ltd as Operator
for and on behalf of the Browse Joint Venture (BJV). We would like to thank the crew of the RV Solander for their professionalism and hard work. We gratefully acknowledge the support from
the Australian Cancer Research Foundation for the Centre for Advanced Cancer Genomics which has made available the Illumina Novaseq6000 systems for the use of Genomics WA. We also thank
Helix Solutions for their support with microsatellite genotyping. This manuscript was written on Whadjuk Noongar _boodja_. We acknowledge the Whadjuk people as traditional owners of this
land and pay respect to their elders past and present. AUTHOR INFORMATION AUTHORS AND AFFILIATIONS * Australian Institute of Marine Science, Indian Ocean Marine Research Centre, Crawley,
Australia L. Thomas, D. Şahin, A. S. Adam, C. M. Grimaldi, N. M. Ryan, S. L. Duffy, J. N. Underwood & J. P. Gilmour * UWA Oceans Institute, The University of Western Australia, Crawley,
Australia L. Thomas, D. Şahin, C. M. Grimaldi, S. L. Duffy, W. J. Kennington & J. P. Gilmour * Centre for Evolutionary Biology, School of Animal Biology, The University of Western
Australia, Perth, Australia W. J. Kennington Authors * L. Thomas View author publications You can also search for this author inPubMed Google Scholar * D. Şahin View author publications You
can also search for this author inPubMed Google Scholar * A. S. Adam View author publications You can also search for this author inPubMed Google Scholar * C. M. Grimaldi View author
publications You can also search for this author inPubMed Google Scholar * N. M. Ryan View author publications You can also search for this author inPubMed Google Scholar * S. L. Duffy View
author publications You can also search for this author inPubMed Google Scholar * J. N. Underwood View author publications You can also search for this author inPubMed Google Scholar * W. J.
Kennington View author publications You can also search for this author inPubMed Google Scholar * J. P. Gilmour View author publications You can also search for this author inPubMed Google
Scholar CONTRIBUTIONS J.G., L.T., and J.U. designed the study, collected samples, analysed the data. D.S., S.D., N.R. and C.G collected samples and analysed data. A.S. and W.J.K. analysed
data. All authors wrote the manuscript. CORRESPONDING AUTHOR Correspondence to L. Thomas. ETHICS DECLARATIONS COMPETING INTERESTS The authors declare no competing interests. PEER REVIEW PEER
REVIEW INFORMATION _Communications Biology_ thanks Chris Langdon, Mikhail Matz and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Primary Handling
Editors: Luciano Matzkin and Luke R. Grinham. A peer review file is available. ADDITIONAL INFORMATION PUBLISHER’S NOTE Springer Nature remains neutral with regard to jurisdictional claims
in published maps and institutional affiliations. SUPPLEMENTARY INFORMATION PEER REVIEW FILE SUPPLEMENTARY INFORMATION DESCRIPTION OF ADDITIONAL SUPPLEMENTARY FILES SUPPLEMENTARY DATA 1
SUPPLEMENTARY DATA 2 SUPPLEMENTARY DATA 3 SUPPLEMENTARY DATA 4 SUPPLEMENTARY DATA 5 REPORTING SUMMARY RIGHTS AND PERMISSIONS OPEN ACCESS This article is licensed under a Creative Commons
Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original
author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the
article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use
is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit
http://creativecommons.org/licenses/by/4.0/. Reprints and permissions ABOUT THIS ARTICLE CITE THIS ARTICLE Thomas, L., Şahin, D., Adam, A.S. _et al._ Resilience to periodic disturbances and
the long-term genetic stability in _Acropora_ coral. _Commun Biol_ 7, 410 (2024). https://doi.org/10.1038/s42003-024-06100-0 Download citation * Received: 29 September 2023 * Accepted: 22
March 2024 * Published: 04 April 2024 * DOI: https://doi.org/10.1038/s42003-024-06100-0 SHARE THIS ARTICLE Anyone you share the following link with will be able to read this content: Get
shareable link Sorry, a shareable link is not currently available for this article. Copy to clipboard Provided by the Springer Nature SharedIt content-sharing initiative