Absence of calcium-sensing receptor basal activity due to inter-subunit disulfide bridges
- Select a language for the TTS:
- UK English Female
- UK English Male
- US English Female
- US English Male
- Australian Female
- Australian Male
- Language selected: (auto detect) - EN
Play all audios:

ABSTRACT G protein-coupled receptors naturally oscillate between inactive and active states, often resulting in receptor constitutive activity with important physiological consequences.
Among the class C G protein-coupled receptors that typically sense amino-acids and their derivatives, the calcium sensing receptor (CaSR) tightly controls blood calcium levels. Its
constitutive activity has not yet been studied. Here, we demonstrate the importance of the inter-subunit disulfide bridges in maintaining the inactive state of CaSR, resulting in
undetectable constitutive activity, unlike the other class C receptors. Deletion of these disulfide bridges results in strong constitutive activity that is abolished by mutations preventing
amino acid binding. It shows that this inter-subunit disulfide link is necessary to limit the agonist effect of amino acids on CaSR. Furthermore, human genetic mutations deleting these
bridges and associated with hypocalcemia result in elevated CaSR constitutive activity. These results highlight the physiological importance of fine tuning the constitutive activity of G
protein-coupled receptors. SIMILAR CONTENT BEING VIEWED BY OTHERS ASYMMETRIC ACTIVATION OF THE CALCIUM-SENSING RECEPTOR HOMODIMER Article 30 June 2021 PROMISCUOUS G-PROTEIN ACTIVATION BY THE
CALCIUM-SENSING RECEPTOR Article 17 April 2024 ALLOSTERIC MODULATION AND G-PROTEIN SELECTIVITY OF THE CA2+-SENSING RECEPTOR Article 07 February 2024 INTRODUCTION Cell surface receptors
generate intracellular signals upon activation by external stimuli. Like all proteins, these receptors exhibit constant conformational dynamics oscillating between inactive and active
states, which has been extensively studied for G protein-coupled receptors (GPCRs)1,2,3,4. Accordingly, receptors that maintain in an active state for sufficiently long periods can signal
without activation by external stimuli5,6,7,8,9. Such constitutive activity can leads to an optimal physiological output under evolutionary pressure, which is biologically significant10.
Indeed, many genetic mutations that have been found to either increase or decrease receptor constitutive activity can be the origin of human diseases6,11,12. For most GPCRs, the constitutive
activity can be easily detected6,13,14,15,16. However, some of them exhibit stable inactive conformations that lead to the undetectable constitutive activity, which permits receptors to
generate signals only upon agonist activation. For instance, α1B adrenergic receptor is well constrained in inactive state by a specific residue in the intracellular loop 317,18. In
addition, the inverse agonist cis-retinal stabilizes the rhodopsin in fully inactive state19. Conversely, some GPCRs exhibit strong constitutive activity that is almost equal to the activity
elicited by agonist stimulation. As an example, the melanocortin-4 receptor, in which endogenous ligands (agouti-related proteins) act as inverse agonists20. The constitutive activity can
be affected by the cellular environment. Indeed, constitutive activity of some metabotropic glutamate receptors (mGluRs), mGluR1 and mGluR5, is naturally inhibited by the long form of the
intracellular partner Homer, but is revealed by the short form Homer1a21. Another example is the constitutive activity of 5-HT6 receptor, which is promoted by its interaction with
neurofibromin22 or Cdk523. GPCR constitutive activity can also be regulated by receptor phosphorylation24, dimerization25, alternative splicing26, or RNA editing27. The discovery of inverse
agonists and antagonists made it possible to identify the physiological significance of GPCR constitutive activity6,10. For example, Homer regulates the constitutive activity of mGluRs,
which is important for synaptic metaplasticity28,29, and the constitutive activity of histamine H3 receptor plays a leading role in regulating the activity of histamine neurons30. The class
C GPCRs that sense amino-acids and their derivatives31 usually exhibit significant constitutive activity21,32,33,34. However, that of the calcium-sensing receptor (CaSR)35 has not been well
studied. CaSR is a multifunctional receptor which plays important physiological role in organs involved or not involved in Ca2+ homeostasis36,37. The fundamental role of CaSR is to enable
parathyroid gland to detect subtle changes in extracellular Ca2+ concentration and to respond by modulating the release of parathyroid hormone (PTH) in the opposite direction38. The result
is Ca2+ normalization through the direct action of PTH, which promotes calcium release from bones and reabsorption by the kidney, in addition to its indirect effects in the gut. In the
parathyroid gland, CaSR signaling activity suppresses PTH secretion, thus preventing the development of hypercalcemia. This role of CaSR is also mediated by cell-autonomous activities in
kidney, bone and thyroid C cells. Consequently, if the receptor remains in a constant state of hyperactivity, it can lead to disturbances in calcium homeostasis and disorders such as
hypocalcemia. Many genetic mutations that lead to loss- or gain-of-function of the CaSR have been identified in patients with metabolic syndromes, such as familial hypocalciuric
hypercalcemia (FHH), neonatal severe primary hyperparathyroidism (NSPHT) or autosomal dominant hypocalcemia (ADH)37,39. In addition, CaSR autoantibodies that induced altered receptor
signaling properties have been identified in acquired hypocalciuric hypercalcemia40. Actually, three commercial drugs can target CaSR as positive allosteric modulator (PAMs).
Etelcalcetide41,42 targets the extracellular domain (ECD) and is applied in the treatment of secondary hyperparathyroidism, and two other drugs can bind to the transmembrane domain and are
used to the treat both primary and secondary hyperparathyroidism43,44. Like other typical class C GPCRs, such as metabotropic glutamate receptors (mGluRs), γ-aminobutyric acid B receptor
(GABABR), GPRC6A and sweet/umami taste receptors, CaSR forms a constitutive dimer45. Each subunit within the dimer is composed of an extracellular Venus flytrap (VFT) domain, a cysteine-rich
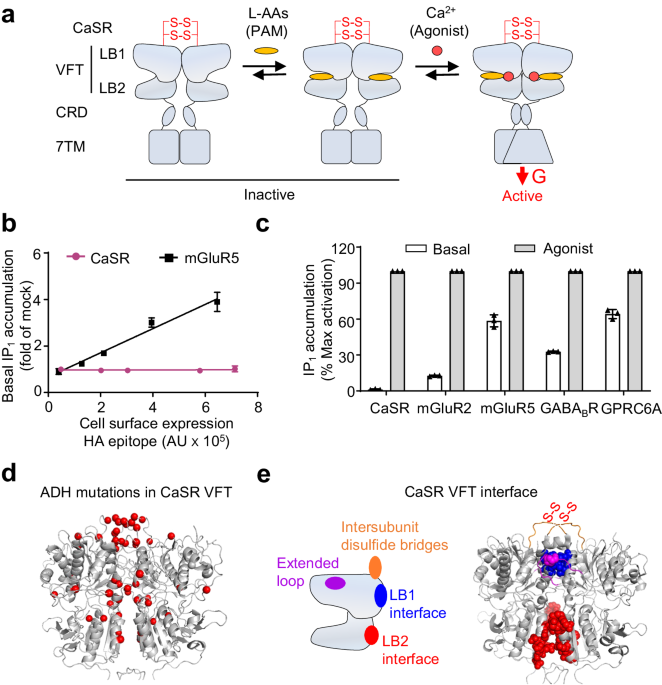
domain (CRD) and a seven-transmembrane (7TM) domain. Among them, the VFT domain is responsible for the binding of calcium ions45 and L-AAs46 (Fig. 1a). Furthermore, the two protomers are
cross-linked by two pairs of disulfide bridges. Recently, the structure of full-length CaSR, both in inactive and active states with L-AAs, which shown to behave as pure-PAMs without direct
agonist effect (Fig. 1a), or with allosteric modulators, including etelcalcetide, have been analyzed47,48,49,50,51,52,53,54. It remains unclear the role of L-AAs in the constitutive activity
of CaSR. Here, we demonstrate that in contrast to other class C GPCRs, CaSR displays no detectable constitutive activity. We reveal that the covalent disulfide bridges between the two
subunits of this dimeric receptor are essential for the limitation of constitutive activity. More importantly, compare with the other class C GPCRs of the mGluR family that possess a single
inter-subunit disulfide bridge, CaSR has two disulfide bridges. Both two disulfide bridges are essential for blocking the constitutive activity and potential agonist effect of ambient L-AAs.
Several mutations of Cys residues that involved in these bridges had been identified in ADH patients. Interestingly, we found that all these mutations could generate constitutive activity,
which revealed the importance of the disulfide bridges in controlling calcemia. These data further illustrate the significant role of the inter-subunit disulfide bridge in class C GPCRs in
controlling their activity, and the importance of an evolutionarily optimized constitutive activity of receptors for proper physiological functions. RESULTS CONSTITUTIVE ACTIVITY OF CASR IS
NOT DETECTABLE We compared the constitutive activity of CaSR with mGluR5 by measuring the inositol phosphate-1 (IP1) accumulation, both of receptors are naturally coupled to the Gq protein.
As reported previously, a high constitutive activity of mGluR5 could be detected55 (Fig. 1b and Supplementary Fig. 1a). In contrast, the IP1 constitutive accumulation in CaSR transfected
HEK-293 cell was unable to be detected in the absence of calcium, even in the group with highest CaSR surface expression (Fig. 1b). Our data indicate that HEK-293 cells do not expressed
endogenous CaSR at a level high enough to be detected (Supplementary Fig. 1b, c). The constitutive activity of mGluR232, GABABR33,34, and GPRC6A56 also can be detected in transfected cells
as described in previous studies (Fig. 1c and Supplementary Fig. 1a). Co-transfected chimeric Gqi9 made it possible to detect the IP1 accumulation in HEK-293 cells that transfected with
mGluR2 and GABABR, for both receptors belong to the Gi/o coupled-receptors57. Altogether, these data showed that the CaSR activity is tightly controlled with a no detectable constitutive
activity toward the Gq pathway. UNDETECTABLE CONSTITUTIVE ACTIVITY OF CASR IS DUE TO THE INTER-SUBUNIT DISULFIDE BRIDGES To identify the molecular basis for the undetectable constitutive
activity in CaSR, we analyzed the numerous missense genetic mutations that have been reported to increase the activity of CaSR in patients with ADH disease39. Many mutations occurred in the
VFT domain (Fig. 1d), especially at the dimer interface that is known to control the structural dynamics and receptor activation2,4,47,58,59,60. Based on the inactive and active structures
of CaSR, there was reasonable to explain that the VFT dimer interface mainly consists of four regions48,51,58 (Fig. 1e). The upper loop contains a pair of cysteine residues (Cys129 and
Cys131) which were responsible for the covalent link between the two protomers, and was highly conserved in CaSR during evolution (Supplementary Fig. 2a). Interestingly, this upper loop
contains a hot spot of ADH mutations39. We therefore investigated the role of the inter-subunit disulfide bridges between the two upper loops (Fig. 2a). These Cys were replaced with Ser
since such natural mutations have been previously reported in ADH61,62. We found that C129S + C131S (CSCS) double mutations resulted in the appearance of monomer band in non-reducing
SDS-PAGE experiments, however, dimeric form of the mutant CSCS still could be detected on the cell membrane (Supplementary Fig. 2b, c). Indeed, dimers and even larger oligomers are observed
for many GPCRs in SDS-PAGE even when they are not covalently linked (i.e. many class A GPCRs). This is likely due to the consequence of unfolding by SDS and mixing the two chains during this
process, resulting in non-covalent dimers that are not sensitive to reducing agents such as DTT63. We further verified this result by bioluminescence resonance energy transfer (BRET) and
obtained a similar conclusion, which suggested that the dimerization of CaSR was maintained by multiple interactions (Supplementary Fig. 2d). Interestingly, both single and double mutants
displayed strong constitutive activity, despite their surface expression was similar to wild-type (WT) receptor (Fig. 2b and Supplementary Fig. 2e). The constitutive activity was positively
correlated with the cell surface expression of the mutated receptors (Fig. 2c) and could be blocked by the CaSR negative allosteric modulator NPS 214364 (Fig. 2d). The mutants also showed a
significant increase in CaCl2 potency compared with WT receptor (Fig. 2b). In addition, in contrast to the WT receptor, without agonist binding, the mutants became more sensitive in the
activation induced by PAMs, NPS R568 or AC265347, both of which bind to the 7TM domain of CaSR65,66 (Fig. 2e). Similar results were obtained by measuring the phosphorylated ERK1/2 (pERK),
these Cys mutants upregulated the basal level of pERK and NPS-2143 treatment inhibited this process (Supplementary Fig. 3a). Altogether, these results showed the absence of inter-subunit
disulfide bonds led to a high basal activity of CaSR. To further validate the correlation between inter-subunit disulfide bond and basal activity, we have re-built one inter-subunit
disulfide bond by changing the Ser132 into Cys in the CSCS construct. This mutant showed a similar dimerization pattern as the WT and single Cys mutants in non-reducing SDS-PAGE experiments
(Supplementary Fig. 3b). Interestingly, the CSCS + S132C construct showed a strong reduction of constitutive activity in both IP1 accumulation (Fig. 2f) and ERK phosphorylation when compared
with the CSCS construct (Supplementary Fig. 3c). Furthermore, addition of this S132C mutation in the CSCS construct could restore the Ca2+ pEC50 similar to the single Cys mutant
(Supplementary Fig. 3d). Of note, an another CaSR construct where both Cys129 and Cys131 were replaced by Ala also showed a similar increase in basal activity, that could be inhibited by
NPS2143 (Supplementary Fig. 3e). These results further indicated that the emergence of the constitutive activity in CaSR is a result of inter-subunit disulfide bridges removal. Taken
together, our data suggested that the inter-subunit disulfide bridges maintain the inactive state of CaSR and were required to restricting the basal activity of receptor. Interestingly, our
data also revealed that the presence of two disulfide bridges were crucial to the strong limitation of the basal activity, since a single mutation of Cys129 or Cys131 led to a moderate
constitutive activity (10–15% of the WT Emax; see Fig. 2b). And the double mutations that totally remove inter-subunit covalent link between VFTs generated a high constitutive activity (30%
of the WT Emax) (Fig. 2g). THE INTER-SUBUNIT DISULFIDE BRIDGE FAVORS THE BASAL ACTIVITY OF MGLURS Since the inter-subunit disulfide bridge is conserved in most class C homodimers, especially
in mGluRs (Fig. 3a), we wondered whether they play a similar role in controlling the basal activation. In contrast to CaSR, mGluRs have only one conserved Cys residue in the upper loop that
could establish single disulfide bridge (Fig. 3b). It was reported that mutated Cys121 to Ala in mGluR2 stabilized the VFT in inactive state4. Consistently, we found that the C121A mutant
has a lower constitutive activity in IP1 accumulation than the WT receptor when co-transfected with Gqi9 (Fig. 3c) and this decrease was not due to the difference in cell surface expression
(Supplementary Fig. 4a). In addition, to mimic the upper loop of CaSR and potentially favor the formation of two inter-subunit disulfide bridges, a second Cys residue was introduced at the
position 119 in mGluR2 equivalent to the first conserved Cys (Cys129) in the CaSR (Fig. 3c and Supplementary Fig. 4a, b). The H119C mutation significantly increased the basal activity of
mGluR2 (Fig. 3c) as well as its potency for glutamate (Fig. 3d). Similar experiments were also performed on mGluR5, which belongs to another group of mGluRs that are naturally coupled to the
Gq—phospholipase C signaling pathways. Similar to the mGluR2 mutants, deletion the inter-subunit disulfide bridge (C129A) in mGluR5 resulted in a lower basal activity, while an additional
Cys (V127C) could increase the basal activity (Fig. 3e and Supplementary Fig. 4c, d) and its potency for glutamate (Fig. 3f). Altogether, our results showed that the inter-subunit disulfide
bridge controls the basal activity of CaSR and mGluRs in distinct ways. REMOVAL OF INTER-SUBUNIT DISULFIDE BRIDGE ENHANCES SIGNALING OUTPUT OF CASR CaSR could respond to diverse ligands and
signals through different G proteins67,68. We investigated whether the CSCS mutant responds differently to these ligands and show preference toward different G proteins. As mentioned above,
the CSCS mutant was more sensitive to Ca2+ in comparison with WT CaSR, and the results from other ligands like Mg2+, neomycin and spermine show a similar effect of these ligands
(Supplementary Fig. 5a). To investigate the influence of this mutant on the activation of different Gq, Gi/o and G13 proteins, we measured the G protein dissociation by using BRET-based
sensors as previously described (Supplementary Fig. 5b)34,69. For all these G proteins, the BRET signal between the Gα and Gβγ subunits was lower in the CSCS group than WT under basal
conditions (Supplementary Fig. 5c–i). Upon agonist stimulation, the BRET signal measured for most G proteins showed increased pEC50 and decreased Emax except G13, which behaved differently
(Supplementary Fig. 5c–j). These results demonstrated that removal of the inter-subunit disulfide bridges in CaSR did not influence G protein selectivity under basal and agonist-induced
conditions. GENETIC MUTATIONS AT CYS129 AND CYS131 FAVOR THE CONSTITUTIVE ACTIVITY OF CASR Among the genetic gain-of-function mutations that are associated with ADH (Fig. 1d), several
mutations occurred at Cys129 or Cys131 (Fig. 4a)61,70. We tested the effect of six natural single mutations on the basal activity of CaSR and found that all these mutants showed a strong
basal activity (Fig. 4b) despite their surface expression was similar to WT receptor (Supplementary Fig. 6a). Furthermore, compared with WT receptor, these mutants showed higher Ca2+ potency
and more sensitive to NRS R568 (Fig. 4c–f). In addition, using the BRET-based G protein sensors, we showed that the mutant C131W, used as an example, has a higher basal activity in
different G protein pathways than WT receptor, and an increased pEC50 and decreased Emax in C131W also been revealed which were in consistence with the results obtained with the CSCS
construct. We also found that the CSCS construct showed higher sensitivity to CaCl2 compared to this C131W mutant except for the GoB pathway (Supplementary Fig. 6b–j). Therefore, the natural
mutations at either C129 or C131 would impair the integrity of the inter-subunit disulfide bridges and led to the basal activation of CaSR. L-AAS BINDING IN CASR IS REQUIRED FOR THE BASAL
ACTIVITY OF THE CSCS CONSTRUCT It was recently reported that ambient L-AAs in the cell culture medium were constitutively bound in the CaSR VFT (Fig. 5a), and act as pure-PAMs during
activation by extracellular Ca2+ 45,47,48,49,50,51,52,53. However, the extracellular concentrations of these L-AAs in cellular assays were difficult to control32,45, even in the absence of
serum. To study the importance of the L-AAs in the basal activity of the CSCS mutant, we constructed several single mutations that are known to prevent L-AAs binding in the VFT binding
pocket of CaSR45,47,49. Interestingly, the mutations abolished the constitutive activity of CSCS mutant (Fig. 5b and Supplementary Fig. 7a). Of note, the gain-of-function mutation E297D71
that was proposed to increase the affinity of L-AAs for CaSR45 had no effect on the basal activity of CSCS (Fig. 5b). Then we washed the cells three times with starvation buffer over 3 h to
remove the L-AAs that had bound to the receptor, and discovered that the basal activity of the mutations were significantly decreased in this group, and it could been recovered by the
addition of L-Trp (Fig. 5c). Elution and rebinding treatments with L-AAs showed that the spontaneous activity of the CSCS mutant was dependent on the intact amino acid binding. Moreover,
this agonistic effect of L-AAs was more significant with the CSCS + E297D construct, which is consistent with the higher sensitivity of the CSCS + E297D construct to L-AAs conferred by the
mutation E297D45. The results suggested that removal of the inter-subunit disulfide bridges altered the pure-PAM effect of L-AAs that become ago-PAMs. We further investigated whether the
binding of ambient L-AAs to both VFTs of the dimer was required to maintain the basal activity. We constructed a ‘controlled’ CaSR homodimer formed by the CaSRC1 and CaSRC2
subunits51,72,73,74. In these constructs, the C-terminus of the CaSR subunits were replaced by the C-terminus from modified GABAB1 (C1) or GABAB2 (C2) subunits, respectively. The dimers that
consisted of the same C-terminus (CaSRC1 + CaSRC1 or CaSRC2 + CaSRC2) hardly reached the cell surface (Fig. 5d and Supplementary Fig. 7b) and responded weakly to the CaCl2 stimulation
(Supplementary Fig. 7c). Then we introduced a S170A single mutation to the CSCSC1-CSCSC2 constructs. This mutation that occurs in CSCSC1 or CSCSC2 alone strongly decreased the basal
activity, and more importantly, when S170A mutation is in both VFTs it abolished the constitutive activity (Fig. 5e and Supplementary Fig. 7d). These results suggested that L-AA binding in
at least one VFT of CSCS was required to generate constitutive activity, and binding in both VFTs could produce the strongest basal activity (Fig. 5f). Altogether, these data showed the
importance of L-AAs induced VFT closure in the basal activity of CSCS. UPPER INTERFACE OF THE VFT IS NOT INVOLVED IN CASR BASAL ACTIVITY In addition to the inter-subunit disulfide bridges,
we wondered whether other regions at the VFT interface could control the constitutive activity of CaSR. Other direct interactions between the two LB1 were the extended loops which could
stabilized the VFT interface through hydrophobic and polar interactions (Fig. 1e and Supplementary Fig. 8a). This unique structure is not conserved in the other class C GPCRs (Supplementary
Fig. 8b). Two loss-of-function genetic mutations in this loop, S53P and P55L39,70 (Supplementary Fig 8a) showed no effect on the basal activity of the receptor (Supplementary Fig. 8c, d).
But both mutants have a lower potency for calcium activation (Supplementary Fig. 8e), which were in line with their classification as inactivating mutations. In addition, we analyzed another
region of the LB1 interface that is conserved in the other class C GPCRs (Supplementary Fig. 8f, g). The two loss-of-function genetic mutations in this region, L159F and L159R75,76, as well
as L108A, L112A, and L156A had no effect on the basal activity of receptor (Supplementary Fig. 8h, i), but decreased the pEC50 of Ca2+ (Supplementary Fig. 8j). On the other hand, the
residue Phe160 located immediately after helix C, which may interact directly with the upper loop (residue Val115) in both the inactive and active states (Supplementary Fig. 8k), was also
mutated. Interestingly, the F160A mutant shows a significant constitutive activity (Supplementary Fig. 8l) and higher Ca2+ potency than the WT (Supplementary Fig. 8j). This is probably due
to an effect on the conformation of the upper loop via the loss of interaction with the side chain of Val115, which could lead to its repositioning. Indeed, Val115 is located between the end
of helix B and the beginning of the upper loop that adopts a helical conformation, a critical position for the relative positioning of helix B and the upper loop (Supplementary Fig. 8k).
Altogether, our data showed that the LB1 interface of the VFTs, apart from the upper loop containing the disulfide bridges, is not involved in CaSR basal activity. NEGATIVE CHARGES IN THE
LOWER INTERFACE OF THE VFT LIMIT CASR ACTIVATION BUT IS NOT INVOLVED IN BASAL ACTIVITY In contrast to mGluRs and other class C GPCRs, the lower interface of the CaSR VFT (LB2) was highly
enriched with negatively charged residues (Fig. 6a, b). Our recent study have ruled out the possibility that this region has a functional binding site for Ca2+ 45. To investigate the effect
of LB2 interface on basal activity, we constructed a mutant where all the 13 negatively charged residues in the acidic patch been replaced by alanine, and we named this mutant 13 A (Fig. 6c,
Supplementary Fig. 9a). The 13 A mutant showed no effect on basal activity when made in the WT receptor, while the introduction of 13 A mutation in the CSCS construct further increased the
basal activity (Fig. 6d). In addition, the similar pEC50 for Ca2+ indicated that introduction of 13 A mutations did not impair the activation of CaSR by Ca2+, both in WT and CSCS constructs
(Fig. 6e), which was consistent to the results obtained with single or multiple mutants in this acidic LB2 patch in previous study45. Interestingly, the 13 A mutations potentiated the
activation of CaSR by NPS R568, both in WT and CSCS constructs (Fig. 6f). Finally, we analyzed the effect of the PAM etelcalcetide, which could form a disulfide bond with the free LB2
Cys48242 and binds to the LB2 interface by forming salt bridges with the patch of negatively charged residues51 (Supplementary Fig. 9b). Etelcalcetide showed no impact on the basal level of
IP1 accumulation in WT receptor, but elevated the basal activity of the CSCS construct (Supplementary Fig. 9c). Moreover, etelcalcetide could increase the Ca2+ potency of WT and CSCS mutant
(Supplementary Fig. 9d), as well as the agonist effect of NPS R568 on both WT and CSCS constructs (Supplementary Fig. 9e). Altogether, our results showed that the negatively charged
interface of LB2 exerted no effect on constitutive activity of WT receptor but could enhance it in the CSCS construct, underscoring the pivotal role of inter-subunit disulfide bonds in
limiting the constitutive activity of CaSR. It also suggests that when both LB2s could come closed, it facilitates the activation of CaSR by PAM binding in the 7TM. DISCUSSION As a special
nutrient-sensing receptor regulated by various ligands, CaSR develops a unique ability to sense the tiny fluctuation of extracellular Ca2+ 77,78. This response is essential for tight control
of calcemia through various mechanisms, including in the parathyroid which controls PTH release, in the bones and kidney among others37. Therefore, the CaSR activity was critical for proper
physiological response. In this study, we showed that inter-subunit disulfide bridges functions as molecular locks to significantly restrict the constitutive activity. They also reduce
receptor sensitivity to the agonists, as well as inhibit the agonism of CaSR by natural and synthetic PAMs, thus setting the Ca2+ potency to an optimal value. Class C GPCRs are complex
GPCRs79, the majority of them have constitutive activity34,55,79,80,81. Their activation mechanism involves several steps, such as the closure and reorientation of VFT, which leads the CRD
closed to each other, followed by a direct interaction between the 7TM domains through TM6 and then a conformational change in one of the 7TM domain60. Consequently, their constitutive
activity can originate from various structural elements. First, as demonstrated in GABABR, it could come from the spontaneous closure of VFT, which is abolished by the competitive
antagonist33,59. Alternatively, it could also originate from any other step following the closure of the VFT such as the CRD and 7TM dimer rearrangement82. As demonstrated in mGluR5, the
constitutive activity could come from the 7TM domain that can reach an active state even if the VFTs remain in their inactive orientation55. GABABR is an additional instance where a genetic
mutation S695I in human GABAB2 7TM could trigger strong constitutive activity, which was associated with epileptic encephalopathy63. As such, several specific structural determinants in the
CaSR could be largely limit or even suppress its constitutive activity. This is in line with the identification of more than one hundred mutations in the ECD and 7TM domains of CaSR in
patients with gain-of-function CaSR associated with ADH39. However, only a few of them, such as N802I in TM6 and A843E in TM7, have been investigated for their constitutive activity83,84. In
the present study, we examined the possible role of the subunit interface at the level of the VFT dimer, as it may regulate the basal activity of CaSR. We show that the inter-subunit
disulfide bridges on the upper loops of LB1 is required to maintain a completely inactive state with low Ca2+ potency in the 2.2–2.6 mM range. This concentration range is optimal for the
physiological function of the receptor in controlling calcemia. We uncovered that all natural mutations occurring at either C129 or C131 could trigger the constitutive activation of CaSR.
Therefore, the integrity of two inter-subunit disulfide bridges is required to abolish constitutive activity of CaSR. This finding might provide an explanation for calcemia induced by
mutations in either C129 or C131. Constitutive internalization and constitutive activity are usually related, and could be blocked by reverse agonists. Previous studies demonstrated that
CaSR undergoes constitutive and agonist induced internalization, both of which were G protein-independent and β-arrestin-dependent at least partially85. Interestingly, a negative allosteric
modulator did not inhibit the constitutive internalization of CaSR, which suggests there is no link between basal activity and constitutive internalization of CaSR. However, it needs to be
examined in more details in future studies. Among the class C GPCRs examined in this study, only CaSR had no detectable constitutive activity, this observation may be attributed to the fact
that CaSR has two inter-subunit bonds. These cysteine residues have been conserved from fish to mammals, illustrating their evolutionary importance and their ability to tightly control a
very low basal activity. In contrast to what was observed for CaSR, the mGluRs and GPRC6A which only have one inter-subunit bridge, mutation of the unique inter-subunit disulfide bond in
mGluR2 reduced the apparent agonist affinity and decreased the stability of the active state4. The distinct roles of inter-subunit disulfide bonds between mGluRs and CaSR may be due to the
different sequence and conformation of the upper loop where the disulfide bridge is located, but also the preceding α-helix B which participates in the dimer interface between the two
protomers. In mGluRs, this helix B adopts a different conformation between the inactive and active states. Interestingly, the loop immediately preceding it contains residues important for
ligand recognition in the VFT binding pocket86. Agonist binding in the VFT can therefore trigger relaxation of the inter-subunit interface by acting on this helix B, which releases its
constraint upon activation. Concerning the upper loop, its structure is often not solved in the class C GPCRs because of its flexibility51,87,88,89. However, in the few structures where this
loop could be observed, its conformation in CaSR and mGluRs is different (Supplementary Fig. 10a–d). Another difference in the activation mechanism between CaSR and mGluRs, which could
affect constitutive activity, is the large amplitude of VFT dimer reorientation observed for mGluRs between inactive and active states. In CaSR, the amplitude of VFT reorientation is not as
large as in the mGluRs. Mutation of the inter-subunit disulfide bridge in mGluRs could limit this VFT reorientation, thus reducing constitutive activity as observed in our study for both
mGluR2 and mGluR5. In brief, it is reasonable to presume that the inter-subunit disulfide bond in the upper loop may affect the receptor’s equilibrium between active and inactive state1,2,
but additional experimental approaches and methods are required to fully investigate the precise mechanisms in the future. Our findings with inter-subunit disulfide bridge mutants provide
insights on the molecular mechanism of CaSR activation. Cryo-EM structures of CaSR confirmed that L-AAs binding stabilizes the closed state of VFT, an intermediate state which is more easily
activated by Ca2+ but does not result in receptor activation49. We uncovered through mutagenesis that the basal activity induced by impairing the inter-subunit disulfide bond was most
probably caused by the ambient L-AAs. And when the integrity of the inter-subunit covalent link was lost, L-AAs occupied only one VFT pocket were sufficient to drive the receptor activation.
Removing the inter-subunit covalent link most likely decreases the activation energy barrier and enabling L-AAs binding to activate the receptor, as well as increasing the agonist potency
of the receptor. This further validates the function of inter-subunit disulfide links in restricting the CaSR activation for better adaptation to complex environmental stimuli, and also
explains why L-AAs have no agonist activity but behave as pure-PAMs45. Our results on the LB2 interface also help to clarify the molecular mechanism of CaSR activation. This region with
highly negatively charged residues is involved in the contact formation between LB2 domains upon the activation of class C GPCRs. Binding of cations, such as Gd3+, can neutralize the
electrostatic repulsion in mGluRs and stabilize receptor active conformation90. The CaSR LB2 interface exhibits significantly higher electrostatic repulsion than mGluRs because it contains
approximately twice as many negatively charged residues45,58. The electrostatic repulsion from this region was proposed to be able to allosterically control CaSR activation58. Our previous
study demonstrated that mutations in this region did not alter the Ca2+ potency and excluded the possibility that this region has a functional binding site for Ca2+ 45. Here we show that
neutralization of negatively charged residues in this region fails to change the Ca2+ potency, but significantly enhanced the agonist effect of PAMs on CaSR and the basal activity of the
CSCS mutant. Therefore, we speculated that the inter-subunit covalent bridges has a stronger importance than the electrostatic repulsion caused by the LB2 interface to stabilize the inactive
state. Disruption of the inter-subunit disulfide bridges could bring the two LB2 lobes into proximity to induce an activation signal. And this activation signal could be further facilitated
by reducing the electrostatic repulsion at the LB2 interface. However, in presence of the inter-subunit disulfide bridges, only reducing the electrostatic repulsion in the receptor did not
alter the calcium potency or basal activity. Overall, inter-subunit disulfide bridges are the main determinant limiting receptor activation by raising the energetic barrier to CaSR
activation. Electrostatic repulsion at the LB2 interface is a secondary determinant that reinforces stabilization of the inactive state. The combination of these two determinants enables
CaSR to accurately detect surrounding complex nutrient signals. Taken together, our study demonstrates that the specific structural framework of VFT provide molecular basis for the unique
activation features of CaSR, permitting for fine tuning of the ligands sensitivity of the receptor. All results allowed us to propose a molecular lock model, which speculates that the
inter-subunit disulfide bonds serve as molecular locks to restrict the receptor’s dynamics and stabilize it in inactive conformations (Fig. 7). Removal of the inter-subunit disulfide bonds
enables L-AAs alone to activate CaSR and elevates the receptor’s agonist potency. Therefore, these findings highlight the importance of optimal constitutive activity of receptors and provide
valuable insights for the design and development of targeted drugs. METHODS MATERIALS Calcium chloride (Cat. 10043-52-4), magnesium chloride (Cat. 7786-30-3), GABA (Cat. A2129), L-glutamate
(Cat. 56-86-0), L-alanine (Cat. 56-41-7), L-trytophan (Cat. 73-22-3), neomycin (Cat. 1405-10-3) and spermine (Cat. 71-44-3) were purchased from Sigma-Aldrich (St. Louis, MO, USA). NPS R-568
(Cat. 3815) and AC265347 (Cat. 6165) were purchased from Tocris Bioscience (Bristol, UK). NPS 2143 (Cat. ab145050) was obtained from Abcam (Cambridge, UK). Etelcalcetide (Cat. HY-P1955A)
was from MedChemExpress (NJ, USA). Lipofectamine 2000 (Cat. 11668019) and Fluo4-AM (Cat. M14206) were supplied by Thermo Fisher Scientific (Waltham, MA, USA). SNAP-Surface 649 (Cat. S9159S)
was from New England Biolabs (Ipswich, MA, USA). IP-One Gq kit (Cat. 62IPAPEB), BRET substrate coelenterazine h (Cat. S2001) and furimazine (Cat. N1120) were from Revvity (Codolet, France)
and Promega (Madison, WI, USA), respectively. PLASMIDS AND TRANSFECTION All the plasmids encoding the receptors CaSR, mGluR2, mGluR5, and GPRC6A have a SNAP-tag before the HA- or Flag-tag.
The pRK5 plasmid encoding wild-type human CaSR, with Flag or HA and SNAP tags inserted after the signal peptide45. We have verified that the Flag and HA-tagged CaSR are similarly expressed
on the cell surface (Supplementary Fig. 11a) and showed they have the same potency and Emax upon calcium stimulation (Supplementary Fig. 11b), under the same transfection conditions. The
pRK5 plasmid encoding the rat mGluR2 or mGluR5, labeled with HA and SNAP inserted just after the signal peptide73,82. The pRK5 plasmid encoding rat GB1, tagged with HA inserted after the
signal peptide, and the rat GB2, tagged with Flag inserted after the signal peptide were kindly provided by the Institut de Génomique Fonctionnelle (Montpellier, France)63. The pRK5 plasmid
encoding wild-type human GPRC6A, with a Flag and SNAP tags inserted after the signal peptide was a gift from Revvity. For BRET saturation experiments, plasmids were generated by inserting
the Rluc or YFP sequence at the C-terminus of Flag-SNAPCaSR. The last 192 residues of CaSR C-terminus were replaced by the sequence coding C1KKXX (C1, the coiled-coil sequence of 47 residues
at the C-terminus of GABAB1) or C2KKXX (C2, 49 residues of GABAB2 coiled-coil region, followed by the endoplasmic reticulum retention signal KKTN) to obtain the plasmids encoding
HA-SNAPCaSRC1 and Flag-SNAPCaSRC2. The mutations for CaSR and mGluRs were generated by site-directed mutagenesis using the QuikChange mutagenesis protocol (Agilent Technologies). All
constructs were verified by DNA sequencing. All experiments were performed in HEK293 (ATCC, CRL-1573, lot: 3449904) cultured in DMEM media (Thermo Fisher Scientific) containing 10% fetal
bovine serum (Thermo Fisher Scientific), 100 units/ml penicillin and 100 μg/ml streptomycin (Thermo Fisher Scientific) at 37 °C and 5% CO2. Cells were transiently transfected either by
electroporation or using lipofectamine 2000 according to the manufacturer protocol (Invitrogen Life Technologies)72. In the electroporation transfection, ten million cells were transfected
with a total of 4 μg plasmids. In the liposome transfection assay, two million cells were transfected with 1 μg of total amounts plasmid of interest. In order to couple mGluR2 and GABABR to
the phospholipase C pathway, the cells were co-transfected with the chimeric G protein Gqi9. For experiments involving mGluRs, the cells were also co-transfected with the glutamate
transporter EAAC1. The ratio of amount plasmids for the constructs expressing the indicated receptor, Gqi9 and EAAC1 is 2:1:1. ENZYME-LINKED IMMUNOSORBENT ASSAY Cell surface and total
expression of the indicated constructs were quantified by an enzyme-linked immunosorbent assay (ELISA). Cell culture medium was removed from 96-well plates 24 h after transfection, then
cells were washed and fixed with 4% paraformaldehyde (Sigma-Aldrich), then blocked with 10% FBS (for the total expression group, 0.1% Triton X-100 (Thermo Fisher Scientific) was required
before this step to increase the permeability of the cell membrane). Flag-tagged constructs were detected with the monoclonal mouse anti-Flag antibody coupled with horseradish peroxidase
(1:20,000; F1804, Sigma-Aldrich). HA-tagged constructs were detected with the rat anti-HA antibody coupled with horseradish peroxidase (1:20,000; 3F10, Roche, Indianapolis, USA). Bound
antibodies were detected by chemoluminescence using a Super Signal substrate (Thermo Fisher Scientific) and a 2103 EnVision Multilabel Plate Reader (Perkin Elmer, Waltham, MA, USA).
INTRACELLULAR CALCIUM MEASUREMENTS Intracellular Ca2+ release was measured in 96-well plates of HEK293 cells with a calcium-sensitive fluorescent dye (Fluo4-AM, Thermo Fisher Scientific)45.
Briefly, 24 h after transfection, cells were washed and pre-incubated with 1 μM Fluo-4 AM in Flex buffer (containing 130 mM NaCl, 5.1 mM KCl, 0.42 mM KH2PO4, 0.32 mM Na2HPO4, 5.27 mM
glucose, 20 mM HEPES, 3.3 mM Na2CO3, 0.1% BSA, 2.5 mM probenecid, pH 7.4) at 37 °C for 1 h. Then, cells were washed once with Flex buffer and 50 μl of this buffer was added into the wells.
And 50 μl of the indicated compounds at 2-fold final concentrations were injected at 20 s during 60 s recording. Fluorescence signals (excitation 485 nm, emission 525 nm) were measured by
using the fluorescence microplate reader Flexstation 3 (Molecular Devices, Sunnyvale, CA, USA). The Ca2+ response is given as the agonist-stimulated fluorescence increase. INOSITOL PHOSPHATE
(IP1) MEASUREMENTS IP1 accumulation was determined in 96-well plates using the IP-One HTRF kit (62IPAPEJ, Revvity) according to the manufacturer’s instructions. Briefly, 24 h after
transfection, cells were washed and incubated with indicated compounds in the Ca2+-free stimulation buffer (containing 10 mM HEPES, 146 mM NaCl, 4.2 mM KCl, 1 g/L glucose, 50 mM LiCl, pH
7.4) at 37 °C for 30 min. The d2-labeled IP1 (IP1-d2) and terbium cryptate-labeled anti-IP1 antibody (Anti-IP1-K) were diluted in lysis buffer (provided by the IP-One HTRF kit) and added in
each well, then the plate was incubated in the dark at room temperature for 1 h before detected by the Multi-mode plate reader (PHERAstar FSX, BMG LABTECH). Results were calculated from the
standard curve in the kit with the fluorescence ratio of IP1-d2 emission at 665 nm over the Anti-IP1-K emission at 620 nm. For the experimental group of L-AAs elution and rebinding
treatments, the Mg2+ and Ca2+ free HBS starvation buffer (containing 10 mM HEPES, 140 mM NaCl, 4 mM KCl, 1 mM KH2PO4, pH 7.4) was used to wash cells for three times during 3 h, then the
cells were incubated with the indicated compounds in the stimulation buffer. BIOLUMINESCENCE RESONANCE ENERGY TRANSFER (BRET) To measure the dimerization of CaSR WT and the indicated
mutants, two million cells were co-transfected with a constant amount of Rluc-tagged receptors (0.2 μg) and an increasing amount of YFP-tagged receptors (0, 0.05, 0.1, 0.2, 0.4, 0.6, 0.8,
and 1 μg) and completed to a total amount of 1.2 μg with the plasmid encoding the pRK5 empty vector. Cell culture medium was removed from 96-well plates 24 h after transfection, then cells
were washed and incubated in HBS buffer (containing 10 mM HEPES, 140 mM NaCl, 4 mM KCl, 1 mM KH2PO4, pH 7.4) at 37 °C. YFP fluorescence was measured before the addition of Rluc substrate
coelenterazine H. Then each well was loaded with 40 μL HBS with coelenterazine H at a final concentration of 5 μM, and the signal was read by a Mithras LB 940 (Berthold Biotechnologies, Bad
Wildbad, Germany) at 37 °C for six cycles. The net BRET was defined as the experimental BRET signal values with the baseline subtracted. For G protein activation experiments, two million
cells were transfected with 0.5 μg receptor plasmid, 0.02 μg the indicated Gα fused to Nluc, 0.2 μg Gβ1 and 0.2 μg Venus-tagged Gγ9 by Lipofectamine 2000. Cell culture medium was removed
from 96-well plates 24 h after transfection, then cells were washed and each well was loaded with 40 μL of the Nluc substrate furimazine at a final concentration of 10 μM. After measuring
the baseline BRET signal, cells were treated with 10 μL of drugs (prepared in HBS at 5-fold final concentration) for an additional 6 cycles. Results were calculated as the ratio of the Venus
emission at 485 nm over the Nluc emission at 530 nm. WESTERN BLOT Twenty-four hours after transfection, cells were washed and incubated with or without the indicated treatments in HBS
buffer at 37 °C for 2 h. Then the cells were lysed with ice-cold lysis buffer (containing 50 mM Tris-HCl pH 7.4, 150 mM NaCl, 1% Triton X-100, 1% sodium deoxycholate, 1% SDS), supplemented
with complete phosphatase inhibitor cocktail (Roche) and incubated on ice for 30 min. Samples were denatured with loading buffer (containing 250 mM Tris-HCl pH 6.8, 50% (v/v) glycerol, 10%
(w/v) SDS, 0.5% (w/v) bromophenol blue) and 10 mM DTT for 10 min at 95 °C, separated by SDS-PAGE (10% w/v), and then transferred to nitrocellulose membranes (Millipore, Billerica, MA, USA)
and washed with blocking buffer (5% nonfat dry milk in Tris-buffered saline and 0.1% Tween 20) for 2 h at 25 °C. The following primary antibodies were used for western blot analysis:
polyclonal anti-p44/42 ERK and anti-phospho-p44/42 ERK antibodies (1:3000; 9101 and 9102, Cell Signaling Technology, Shanghai, China), monoclonal Flag antibody (1:1000; KM8002, Sungene
Biotech, Tianjin Province, China) and polyclonal anti-α-tubulin antibody (1:3000; KM9007, Sungene Biotech, Tianjin Province, China). The primary antibodies were incubated overnight at 4 °C,
followed by incubation with the DyLight 800 4 X PEG-conjugated secondary antibody (1:20,000; 5151 and 5257, Cell Signaling Technology, Shanghai, China) for 2 h at 25 °C. The specific protein
bands were visualized by Odyssey CLx imager (LI-COR Bioscience, Lincoln, NE, USA). The density of bands was measured by ImageJ software (Bethesda, MD, USA). FLUORESCENT-LABELED BLOT
EXPERIMENTS The dimerization pattern of CaSR mutants were measured by fluorescent-labeled blot57,63. Briefly, 24 h after transfection, adherent HEK293 cells in 12-well plates were incubated
with 100 nM SNAP-Surface 649 in culture medium in the dark at 37 °C for 1 h, and then lysed with lysis buffer (containing 50 mM Tris-HCl pH 7.4, 150 mM NaCl, 1% NP-40, 0.5% sodium
deoxycholate, 0.1% SDS) for 1 h at 4 °C. After centrifugation at 12,000 × _g_ for 30 min at 4 °C, the supernatants were mixed with loading buffer (containing 250 mM Tris-HCl pH 6.8, 50%
(v/v) glycerol, 10% (w/v) SDS, 0.5% (w/v) bromophenol blue) at 37 °C for 10 min and separated by SDS-PAGE (8% w/v). In reducing conditions, samples were treated with 100 mM DTT in loading
buffer for 10 min before loading the samples. Proteins were transferred to nitrocellulose membranes (Millipore). The specific protein bands were visualized by Odyssey CLx imager (LI-COR
Bioscience). REVERSE TRANSCRIPTION PCR AND REAL-TIME PCR Total RNA of mock and CaSR-overexpressed cells were extracted using Trizol reagent (Invitrogen, Carlsbad, USA) and reverse
transcribed into cDNA using the Superscript first-strand synthesis system (Invitrogen). _GAPDH_ and _CASR_ mRNA expressions were examined. The primers used were as follows: _GAPDH_
(NM_001256799), 5′-tcaccagggctgcttttaacc-3′ and 5′-gacaagcttcccgttctcag-3′; _CASR_ (NM_000388), 5′-ccctctacgattgctgtggt-3′ and 5′-agtctgctggaggaggcata-3′. Real-time PCR was conducted using
the SYBR Green (Vazyme Biotechnology, Nanjing, China) and the StepOne plus Real-time PCR System (Thermo Fisher Scientific). The threshold was set according to the exponential phase of
products, and the cycle threshold (CT) value for samples was determined. The resulting data were analyzed with the comparative CT method for relative gene-expression quantification against
_GAPDH_. MOLECULAR MODELING The structures of the receptors involved were analyzed and displayed with PyMol software (Palo Alto, CA, USA). The molecular models were generated with PyMol
software based on the structures from the PDB (7DTV, 7DTW, 5FBK, 7M3E, 7M3G, 7M3J and 7M3F for CaSR, 7EPA and 7EPB for mGluR2, 7FD8 for mGluR5). The sequence of CaSR between different
species or the human CaSR and different rat mGluR subtypes were aligned with Clustal Omega and displayed by ESPript 391. STATISTICS AND REPRODUCIBILITY Data were analyzed with Prism 7
software (GraphPad Software, San Diego, CA, USA). Concentration-responses curve parameters were derived using a four parameters non-linear regression equation. Unless stated otherwise, data
shown in the figures represent the mean ± S.E.M. of at least three independent experiments performed in triplicates. Statistical differences were determined by GraphPad Prism using one-way
ANOVA with a Dunnett’s multiple comparison test, two-way ANOVA with Sidak’s multiple comparisons test or t test. _P_ < 0.05 was considered to be statistically significant. DATA
AVAILABILITY Data supporting the findings of this manuscript are available from the corresponding authors upon reasonable request. The source data underlying the graphs in the manuscript are
shown in Supplementary Data. The uncropped unedited blot/gel images were shown in Supplementary Figs. 12 and 13. REFERENCES * Olofsson, L. et al. Fine tuning of sub-millisecond
conformational dynamics controls metabotropic glutamate receptors agonist efficacy. _Nat. Commun._ 5, 5206 (2014). Article CAS PubMed Google Scholar * Lecat-Guillet, N. et al. Concerted
conformational changes control metabotropic glutamate receptor activity. _Sci. Adv._ 9, eadf1378 (2023). Article CAS PubMed PubMed Central Google Scholar * Gregorio, G. G. et al.
Single-molecule analysis of ligand efficacy in beta(2)AR-G-protein activation. _Nature_ 547, 68–73 (2017). Article CAS PubMed PubMed Central Google Scholar * Levitz, J. et al. Mechanism
of assembly and cooperativity of homomeric and heteromeric metabotropic glutamate receptors. _Neuron_ 92, 143–159 (2016). Article CAS PubMed PubMed Central Google Scholar * Berg, K. A.
& Clarke, W. P. Making sense of pharmacology: inverse agonism and functional selectivity. _Int. J. Neuropsychopharmacol._ 21, 962–977 (2018). Article CAS PubMed PubMed Central
Google Scholar * Seifert, R. & Wenzel-Seifert, K. Constitutive activity of G-protein-coupled receptors: cause of disease and common property of wild-type receptors. _Naunyn
Schmiedebergs Arch. Pharm._ 366, 381–416 (2002). Article CAS Google Scholar * Schutz, W. & Freissmuth, M. Reverse intrinsic activity of antagonists on G protein-coupled receptors.
_Trends Pharm. Sci._ 13, 376–380 (1992). Article CAS PubMed Google Scholar * Lefkowitz, R. J., Cotecchia, S., Samama, P. & Costa, T. Constitutive activity of receptors coupled to
guanine nucleotide regulatory proteins. _Trends Pharm. Sci._ 14, 303–307 (1993). Article CAS PubMed Google Scholar * Smit, M. J. et al. Pharmacogenomic and structural analysis of
constitutive g protein-coupled receptor activity. _Annu. Rev. Pharm. Toxicol._ 47, 53–87 (2007). Article CAS Google Scholar * Meye, F. J., Ramakers, G. M. & Adan, R. A. The vital role
of constitutive GPCR activity in the mesolimbic dopamine system. _Transl. Psychiatry_ 4, e361 (2014). Article CAS PubMed PubMed Central Google Scholar * Guo, G. et al.
Ligand-Independent EGFR Signaling. _Cancer Res._ 75, 3436–3441 (2015). Article CAS PubMed PubMed Central Google Scholar * Tao, Y. X. Constitutive activation of G protein-coupled
receptors and diseases: insights into mechanisms of activation and therapeutics. _Pharm. Ther._ 120, 129–148 (2008). Article CAS Google Scholar * Lu, S., Jang, W., Inoue, A. &
Lambert, N. A. Constitutive G protein coupling profiles of understudied orphan GPCRs. _PLoS ONE_ 16, e0247743 (2021). Article CAS PubMed PubMed Central Google Scholar * Xu, L. et al.
Cryo-EM structure of constitutively active human Frizzled 7 in complex with heterotrimeric G(s). _Cell Res._ 31, 1311–1314 (2021). Article CAS PubMed PubMed Central Google Scholar *
Zhang, B., Albaker, A., Plouffe, B., Lefebvre, C. & Tiberi, M. Constitutive activities and inverse agonism in dopamine receptors. _Adv. Pharm._ 70, 175–214 (2014). Article CAS Google
Scholar * Wilde, C. et al. The constitutive activity of the adhesion GPCR GPR114/ADGRG5 is mediated by its tethered agonist. _FASEB J._ 30, 666–673 (2016). Article CAS PubMed Google
Scholar * Cotecchia, S. Constitutive activity and inverse agonism at the alpha1adrenoceptors. _Biochem Pharm._ 73, 1076–1083 (2007). Article CAS PubMed Google Scholar * Kjelsberg, M.
A., Cotecchia, S., Ostrowski, J., Caron, M. G. & Lefkowitz, R. J. Constitutive activation of the alpha 1B-adrenergic receptor by all amino acid substitutions at a single site. Evidence
for a region which constrains receptor activation. _J. Biol. Chem._ 267, 1430–1433 (1992). Article CAS PubMed Google Scholar * Fanelli, F., Felline, A. & Marigo, V. Structural
aspects of rod opsin and their implication in genetic diseases. _Pflug. Arch._ 473, 1339–1359 (2021). Article CAS Google Scholar * Tao, Y. X. Constitutive activity in melanocortin-4
receptor: biased signaling of inverse agonists. _Adv. Pharm._ 70, 135–154 (2014). Article CAS Google Scholar * Ango, F. et al. Agonist-independent activation of metabotropic glutamate
receptors by the intracellular protein Homer. _Nature_ 411, 962–965 (2001). Article CAS PubMed Google Scholar * Deraredj Nadim, W. et al. Physical interaction between neurofibromin and
serotonin 5-HT6 receptor promotes receptor constitutive activity. _Proc. Natl Acad. Sci. USA_ 113, 12310–12315 (2016). Article PubMed PubMed Central Google Scholar * Duhr, F. et al. Cdk5
induces constitutive activation of 5-HT6 receptors to promote neurite growth. _Nat. Chem. Biol._ 10, 590–597 (2014). Article CAS PubMed Google Scholar * Murat, S. et al. 5-HT2A
receptor-dependent phosphorylation of mGlu2 receptor at Serine 843 promotes mGlu2 receptor-operated Gi/o signaling. _Mol. Psychiatry_ 24, 1610–1626 (2019). Article CAS PubMed Google
Scholar * Kwon, Y. et al. Dimerization of beta2-adrenergic receptor is responsible for the constitutive activity subjected to inverse agonism. _Cell Chem. Biol._ 29, 1532–1540.e5 (2022).
Article CAS PubMed Google Scholar * De Maeyer, J. H., Aerssens, J., Verhasselt, P. & Lefebvre, R. A. Alternative splicing and exon duplication generates 10 unique porcine 5-HT 4
receptor splice variants including a functional homofusion variant. _Physiol. Genomics_ 34, 22–33 (2008). Article PubMed Google Scholar * Labasque, M. et al. Constitutive activity of
serotonin 2C receptors at G protein-independent signaling: modulation by RNA editing and antidepressants. _Mol. Pharm._ 78, 818–826 (2010). Article CAS Google Scholar * Hu, J. H. et al.
Homeostatic scaling requires group I mGluR activation mediated by Homer1a. _Neuron_ 68, 1128–1142 (2010). Article CAS PubMed PubMed Central Google Scholar * Bockaert, J., Perroy, J.
& Ango, F. The complex formed by group I metabotropic glutamate receptor (mGluR) and homer1a plays a central role in metaplasticity and homeostatic synaptic scaling. _J. Neurosci._ 41,
5567–5578 (2021). Article CAS PubMed PubMed Central Google Scholar * Arrang, J. M., Morisset, S. & Gbahou, F. Constitutive activity of the histamine H3 receptor. _Trends Pharm.
Sci._ 28, 350–357 (2007). Article CAS PubMed Google Scholar * Kniazeff, J., Prezeau, L., Rondard, P., Pin, J. P. & Goudet, C. Dimers and beyond: The functional puzzles of class C
GPCRs. _Pharm. Ther._ 130, 9–25 (2011). Article CAS Google Scholar * Doornbos, M. L. J. et al. Constitutive activity of the metabotropic glutamate receptor 2 explored with a whole-cell
label-free biosensor. _Biochem. Pharm._ 152, 201–210 (2018). Article CAS PubMed Google Scholar * Grunewald, S. et al. Importance of the gamma-aminobutyric acid(B) receptor C-termini for
G-protein coupling. _Mol. Pharm._ 61, 1070–1080 (2002). Article CAS Google Scholar * Xu, C. et al. Specific pharmacological and G(i/o) protein responses of some native GPCRs in neurons.
_Nat. Commun._ 15, 1990 (2024). Article CAS PubMed PubMed Central Google Scholar * Leach, K. et al. International Union of Basic and Clinical Pharmacology. CVIII. Calcium-sensing
receptor nomenclature, pharmacology, and function. _Pharm. Rev._ 72, 558–604 (2020). Article CAS PubMed Google Scholar * Husted, A. S., Trauelsen, M., Rudenko, O., Hjorth, S. A. &
Schwartz, T. W. GPCR-mediated signaling of metabolites. _Cell Metab._ 25, 777–796 (2017). Article CAS PubMed Google Scholar * Hannan, F. M., Kallay, E., Chang, W., Brandi, M. L. &
Thakker, R. V. The calcium-sensing receptor in physiology and in calcitropic and noncalcitropic diseases. _Nat. Rev. Endocrinol._ 15, 33–51 (2018). Article PubMed PubMed Central Google
Scholar * Brown, E. M. et al. Cloning and characterization of an extracellular Ca(2+)-sensing receptor from bovine parathyroid. _Nature_ 366, 575–580 (1993). Article CAS PubMed Google
Scholar * Gorvin, C. M. Molecular and clinical insights from studies of calcium-sensing receptor mutations. _J. Mol. Endocrinol._ 63, R1–R16 (2019). Article CAS PubMed Google Scholar *
Makita, N. et al. Successful prednisolone or calcimimetic treatment of acquired hypocalciuric hypercalcemia caused by biased allosteric CaSR autoantibodies. _JCI Insight_ 7, e156742 (2022).
* Walter, S. et al. Pharmacology of AMG 416 (Velcalcetide), a novel peptide agonist of the calcium-sensing receptor, for the treatment of secondary hyperparathyroidism in hemodialysis
patients. _J. Pharm. Exp. Ther._ 346, 229–240 (2013). Article CAS Google Scholar * Alexander, S. T. et al. Critical cysteine residues in both the calcium-sensing receptor and the
allosteric activator AMG 416 underlie the mechanism of action. _Mol. Pharm._ 88, 853–865 (2015). Article CAS Google Scholar * Nemeth, E. F. et al. Pharmacodynamics of the type II
calcimimetic compound cinacalcet HCl. _J. Pharm. Exp. Ther._ 308, 627–635 (2004). Article CAS Google Scholar * Kawata, T. et al. A novel calcimimetic agent, evocalcet (MT-4580/KHK7580),
suppresses the parathyroid cell function with little effect on the gastrointestinal tract or CYP isozymes in vivo and in vitro. _PLoS ONE_ 13, e0195316 (2018). Article PubMed PubMed
Central Google Scholar * Liu, H. et al. Illuminating the allosteric modulation of the calcium-sensing receptor. _Proc. Natl Acad. Sci. USA_ 117, 21711–21722 (2020). Article CAS PubMed
PubMed Central Google Scholar * Conigrave, A. D., Quinn, S. J. & Brown, E. M. L-amino acid sensing by the extracellular Ca2+-sensing receptor. _Proc. Natl Acad. Sci. USA_ 97, 4814–4819
(2000). Article CAS PubMed PubMed Central Google Scholar * Geng, Y. et al. Structural mechanism of ligand activation in human calcium-sensing receptor. _Elife_ 5, e13662 (2016). *
Zhang, C. et al. Structural basis for regulation of human calcium-sensing receptor by magnesium ions and an unexpected tryptophan derivative co-agonist. _Sci. Adv._ 2, e1600241 (2016).
Article PubMed PubMed Central Google Scholar * Ling, S. et al. Structural mechanism of cooperative activation of the human calcium-sensing receptor by Ca(2+) ions and L-tryptophan. _Cell
Res_ 31, 383–394 (2021). Article CAS PubMed PubMed Central Google Scholar * Chen, X. et al. Structural insights into the activation of human calcium-sensing receptor. _Elife_ 10,
e68578 (2021). * Gao, Y. et al. Asymmetric activation of the calcium-sensing receptor homodimer. _Nature_ 595, 455–459 (2021). Article CAS PubMed PubMed Central Google Scholar * Park,
J. et al. Symmetric activation and modulation of the human calcium-sensing receptor. _Proc. Natl Acad. Sci. USA_ 118, e2115849118 (2021). Article CAS PubMed PubMed Central Google Scholar
* Wen, T. et al. Structural basis for activation and allosteric modulation of full-length calcium-sensing receptor. _Sci. Adv._ 7, eabg1483 (2021). Article CAS PubMed PubMed Central
Google Scholar * Gorvin, C. M. Recent advances in calcium-sensing receptor structures and signaling pathways. _Prog. Mol. Biol. Transl. Sci._ 195, 121–135 (2023). Article CAS PubMed
Google Scholar * Goudet, C. et al. Heptahelical domain of metabotropic glutamate receptor 5 behaves like rhodopsin-like receptors. _Proc. Natl Acad. Sci. USA_ 101, 378–783 (2004). Article
CAS PubMed Google Scholar * Jorgensen, C. V. & Brauner-Osborne, H. Pharmacology and physiological function of the orphan GPRC6A receptor. _Basic Clin. Pharm. Toxicol._ 126, 77–87
(2020). Article CAS Google Scholar * Xue, L. et al. Major ligand-induced rearrangement of the heptahelical domain interface in a GPCR dimer. _Nat. Chem. Biol._ 11, 134–140 (2015). Article
CAS PubMed Google Scholar * Schamber, M. R. & Vafabakhsh, R. Mechanism of sensitivity modulation in the calcium-sensing receptor via electrostatic tuning. _Nat. Commun._ 13, 2194
(2022). Article CAS PubMed PubMed Central Google Scholar * Lecat-Guillet, N. et al. FRET-based sensors unravel activation and allosteric modulation of the GABA(B) receptor. _Cell Chem.
Biol._ 24, 360–370 (2017). Article CAS PubMed Google Scholar * Rondard, P. & Pin, J. P. Dynamics and modulation of metabotropic glutamate receptors. _Curr. Opin. Pharm._ 20, 95–101
(2015). Article CAS Google Scholar * Dong, B. et al. Calcilytic ameliorates abnormalities of mutant calcium-sensing receptor (CaSR) knock-in mice mimicking autosomal dominant hypocalcemia
(ADH). _J. Bone Min. Res_. 30, 1980–1993 (2015). Article CAS Google Scholar * Thim, S. B., Birkebaek, N. H., Nissen, P. H. & Host, C. Activating calcium-sensing receptor gene
variants in children: a case study of infant hypocalcaemia and literature review. _Acta Paediatr._ 103, 1117–1125 (2014). Article CAS PubMed Google Scholar * Xue, L. et al. Rearrangement
of the transmembrane domain interfaces associated with the activation of a GPCR hetero-oligomer. _Nat. Commun._ 10, 2765 (2019). Article PubMed PubMed Central Google Scholar * Jacobsen,
S. E., Gether, U. & Brauner-Osborne, H. Investigating the molecular mechanism of positive and negative allosteric modulators in the calcium-sensing receptor dimer. _Sci. Rep._ 7, 46355
(2017). Article CAS PubMed PubMed Central Google Scholar * Keller, A. N. et al. Identification of global and ligand-specific calcium sensing receptor activation mechanisms. _Mol.
Pharm._ 93, 619–630 (2018). Article CAS Google Scholar * Leach, K. et al. Towards a structural understanding of allosteric drugs at the human calcium-sensing receptor. _Cell Res._ 26,
574–592 (2016). Article CAS PubMed PubMed Central Google Scholar * Gerbino, A. & Colella, M. The different facets of extracellular calcium sensors: old and new concepts in
calcium-sensing receptor signalling and pharmacology. _Int J. Mol. Sci._ 19, 999 (2018). Article PubMed PubMed Central Google Scholar * Abid, H. A., Inoue, A. & Gorvin, C. M.
Heterogeneity of G protein activation by the calcium-sensing receptor. _J. Mol. Endocrinol._ 67, 41–53 (2021). Article CAS PubMed PubMed Central Google Scholar * Schihada, H., Shekhani,
R. & Schulte, G. Quantitative assessment of constitutive G protein-coupled receptor activity with BRET-based G protein biosensors. _Sci. Signal_ 14, eabf1653 (2021). Article CAS
PubMed Google Scholar * Hendy, G. N., Guarnieri, V. & Canaff, L. Calcium-sensing receptor and associated diseases. _Prog. Mol. Biol. Transl. Sci._ 89, 31–95 (2009). Article CAS
PubMed Google Scholar * Silve, C. et al. Delineating a Ca2+ binding pocket within the venus flytrap module of the human calcium-sensing receptor. _J. Biol. Chem._ 280, 37917–37923 (2005).
Article CAS PubMed Google Scholar * Doumazane, E. et al. A new approach to analyze cell surface protein complexes reveals specific heterodimeric metabotropic glutamate receptors. _FASEB
J._ 25, 66–77 (2011). Article CAS PubMed Google Scholar * Liu, J. et al. Allosteric control of an asymmetric transduction in a G protein-coupled receptor heterodimer. _Elife_ 6, e26985
(2017). Article PubMed PubMed Central Google Scholar * Goolam, M. A. et al. Cell surface calcium-sensing receptor heterodimers: mutant gene dosage affects Ca(2+) sensing but not G
protein interaction. _J. Bone Min. Res._ 37, 1787–1807 (2022). Article CAS Google Scholar * Ho, J. et al. Kabuki syndrome and Crohn disease in a child with familial hypocalciuric
hypercalcemia. _J. Pediatr. Endocrinol. Metab._ 23, 975–979 (2010). Article CAS PubMed Google Scholar * Glaudo, M. et al. Heterozygous inactivating CaSR mutations causing neonatal
hyperparathyroidism: function, inheritance and phenotype. _Eur. J. Endocrinol._ 175, 421–431 (2016). Article CAS PubMed Google Scholar * Brown, E. M. & MacLeod, R. J. Extracellular
calcium sensing and extracellular calcium signaling. _Physiol. Rev._ 81, 239–297 (2001). Article CAS PubMed Google Scholar * Centeno, P. P. et al. Phosphate acts directly on the
calcium-sensing receptor to stimulate parathyroid hormone secretion. _Nat. Commun._ 10, 4693 (2019). Article PubMed PubMed Central Google Scholar * Pin, J. P. & Bettler, B.
Organization and functions of mGlu and GABA(B) receptor complexes. _Nature_ 540, 60–68 (2016). Article CAS PubMed Google Scholar * Tora, A. S. et al. Chloride ions stabilize the
glutamate-induced active state of the metabotropic glutamate receptor 3. _Neuropharmacology_ 140, 275–286 (2018). Article CAS PubMed Google Scholar * Liu, L. et al. Allosteric ligands
control the activation of a class C GPCR heterodimer by acting at the transmembrane interface. _Elife_ 10, e70188 (2021). Article CAS PubMed PubMed Central Google Scholar * Huang, S. L.
et al. Interdomain movements in metabotropic glutamate receptor activation. _Proc. Natl Acad. Sci. USA_ 108, 15480–15485 (2011). Article CAS PubMed PubMed Central Google Scholar *
Lia-Baldini, A.-S. et al. Two novel mutations of the calcium-sensing receptor gene affecting the same amino acid position lead to opposite phenotypes and reveal the importance of p.N802 on
receptor activity. _Eur. J. Endocrinol._ 168, K27–K34 (2013). Article CAS PubMed Google Scholar * Zhao, X. M., Hauache, O., Goldsmith, P. K., Collins, R. & Spiegel, A. M. A missense
mutation in the seventh transmembrane domain constitutively activates the human Ca2+ receptor. _FEBS Lett._ 448, 180–184 (1999). Article CAS PubMed Google Scholar * Mos, I., Jacobsen, S.
E., Foster, S. R. & Bräuner-Osborne, H. Calcium-sensing receptor internalization is β-arrestin-dependent and modulated by allosteric ligands. _Mol. Pharm._ 96, 463–474 (2019). Article
CAS Google Scholar * Koehl, A. et al. Structural insights into the activation of metabotropic glutamate receptors. _Nature_ 566, 79–84 (2019). Article CAS PubMed PubMed Central Google
Scholar * Koehl, A. et al. Author Correction: Structural insights into the activation of metabotropic glutamate receptors. _Nature_ 567, E10 (2019). Article CAS PubMed Google Scholar *
Du, J. et al. Structures of human mGlu2 and mGlu7 homo- and heterodimers. _Nature_ 594, 589–593 (2021). Article CAS PubMed Google Scholar * Seven, A. B. et al. G-protein activation by a
metabotropic glutamate receptor. _Nature_ 595, 450–454 (2021). Article CAS PubMed PubMed Central Google Scholar * Tsuchiya, D., Kunishima, N., Kamiya, N., Jingami, H. & Morikawa, K.
Structural views of the ligand-binding cores of a metabotropic glutamate receptor complexed with an antagonist and both glutamate and Gd3+. _Proc. Natl Acad. Sci. USA_ 99, 2660–2665 (2002).
Article CAS PubMed PubMed Central Google Scholar * Sievers, F. & Higgins, D. G. Clustal Omega for making accurate alignments of many protein sequences. _Protein Sci._ 27, 135–145
(2018). Article CAS PubMed Google Scholar Download references ACKNOWLEDGEMENTS This study was supported by grants from National Key R&D Program of China (grant numbers 2022YFE0116600
and 2021ZD0203302 to J.L.), National Natural Science Foundation of China (NSFC) (grant numbers 32330049 and 82320108021 to J.L., 31070737 and 31371423 to P.Y.), Key Technologies R&D
Program of Guangdong Province (grant number 2010A080813001 to J.L.). P.R. was supported by Fondation pour la Recherche Médicale (EQU202303016470), the Institut National de la Santé et de la
Recherche Médicale (INSERM; International Research Program «Brain Signal») and the Franco-Chinese Joint Scientific and Technological Commission (CoMix) from the French Embassy in China.
AUTHOR INFORMATION AUTHORS AND AFFILIATIONS * Cellular Signaling Laboratory, International Research Center for Sensory Biology and Technology of MOST, Key Laboratory of Molecular Biophysics
of MOE, and College of Life Science and Technology, Huazhong University of Science and Technology, Wuhan, Hubei, China Shumin Ma, Xueliang Yin, Ping Yi & Jianfeng Liu * Institut de
Génomique Fonctionnelle (IGF), Université de Montpellier, CNRS, INSERM, Montpellier, Cedex 5, France Jean-Philippe Pin & Philippe Rondard Authors * Shumin Ma View author publications You
can also search for this author inPubMed Google Scholar * Xueliang Yin View author publications You can also search for this author inPubMed Google Scholar * Jean-Philippe Pin View author
publications You can also search for this author inPubMed Google Scholar * Philippe Rondard View author publications You can also search for this author inPubMed Google Scholar * Ping Yi
View author publications You can also search for this author inPubMed Google Scholar * Jianfeng Liu View author publications You can also search for this author inPubMed Google Scholar
CONTRIBUTIONS S.M. and P.Y. designed research; S.M. and X.Y. performed research; S.M., P.Y., J-P.P., and P.R. analyzed data; and S.M., P.Y., J-P.P., P.R., and J.L. wrote the paper.
CORRESPONDING AUTHORS Correspondence to Philippe Rondard, Ping Yi or Jianfeng Liu. ETHICS DECLARATIONS COMPETING INTERESTS Philippe Rondard and Jean-Philippe Pin are involved in a
collaborative team between the CNRS and Revvity (IGF, Montpellier). All other authors declare no competing interests. PEER REVIEW PEER REVIEW INFORMATION _Communications Biology_ thanks
Caroline Gorvin, Jan Voss, and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Primary Handling Editors: Isabelle Lucet and David Favero. ADDITIONAL
INFORMATION PUBLISHER’S NOTE Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations. SUPPLEMENTARY INFORMATION SUPPLEMENTARY
INFORMATION DESCRIPTION OF ADDITIONAL SUPPLEMENTARY FILES SUPPLEMENTARY DATA REPORTING-SUMMARY RIGHTS AND PERMISSIONS OPEN ACCESS This article is licensed under a Creative Commons
Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original
author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the
article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use
is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit
http://creativecommons.org/licenses/by/4.0/. Reprints and permissions ABOUT THIS ARTICLE CITE THIS ARTICLE Ma, S., Yin, X., Pin, JP. _et al._ Absence of calcium-sensing receptor basal
activity due to inter-subunit disulfide bridges. _Commun Biol_ 7, 501 (2024). https://doi.org/10.1038/s42003-024-06189-3 Download citation * Received: 21 August 2023 * Accepted: 12 April
2024 * Published: 25 April 2024 * DOI: https://doi.org/10.1038/s42003-024-06189-3 SHARE THIS ARTICLE Anyone you share the following link with will be able to read this content: Get shareable
link Sorry, a shareable link is not currently available for this article. Copy to clipboard Provided by the Springer Nature SharedIt content-sharing initiative