Thyroid hormone status defines brown adipose tissue activity and browning of white adipose tissues in mice
- Select a language for the TTS:
- UK English Female
- UK English Male
- US English Female
- US English Male
- Australian Female
- Australian Male
- Language selected: (auto detect) - EN
Play all audios:

ABSTRACT The present study aimed to determine the effect of thyroid hormone dysfunction on brown adipose tissue activity and white adipose tissue browning in mice. Twenty randomized female
C57BL/6NTac mice per treatment group housed at room temperature were rendered hypothyroid or hyperthyroid. _In-vivo_ small animal 18F-FDG PET/MRI was performed to determine the effects of
hypo- and hyperthyroidism on BAT mass and BAT activity. _Ex-vivo_14C-acetate loading assay and assessment of thermogenic gene and protein expression permitted analysis of oxidative and
thermogenic capacities of WAT and BAT of eu-, hyper and hypothyroid mice. 18F-FDG PET/MRI revealed a lack of brown adipose tissue activity in hypothyroid mice, whereas hyperthyroid mice
displayed increased BAT mass alongside enhanced 18F-FDG uptake. In white adipose tissue of both, hyper- and hypothyroid mice, we found a significant induction of thermogenic genes together
with multilocular adipocytes expressing UCP1. Taken together, these results suggest that both the hyperthyroid and hypothyroid state stimulate WAT thermogenesis most likely as a consequence
of enhanced adrenergic signaling or compensation for impaired BAT function, respectively. SIMILAR CONTENT BEING VIEWED BY OTHERS TRIIODOTHYRONINE (T3) PROMOTES BROWNING OF WHITE ADIPOSE
THROUGH INHIBITION OF THE PI3K/AKT SIGNALLING PATHWAY Article Open access 02 September 2024 BONE MARROW ADIPOSE TISSUE IS A UNIQUE ADIPOSE SUBTYPE WITH DISTINCT ROLES IN GLUCOSE HOMEOSTASIS
Article Open access 18 June 2020 TRIIODOTHYRONINE (T3) PROMOTES BROWN FAT HYPERPLASIA VIA THYROID HORMONE RECEPTOR Α MEDIATED ADIPOCYTE PROGENITOR CELL PROLIFERATION Article Open access 13
June 2022 INTRODUCTION Thyroid hormones (TH) are intimately involved in the regulation of energy metabolism. Administration of TH to mammals leads to an increase in basal metabolic rate and
thermogenesis1. The important role of TH in the regulation of body temperature homeostasis is perhaps best reflected by the cold or heat intolerance of animals and humans with hypothyroidism
or hyperthyroidism, respectively2. Under conditions of thermoneutrality, core body temperature can be maintained in the absence of any thermoregulatory process (e.g. shivering). However,
under conditions of cold exposure adaptive thermogenesis occurs through a compensatory increase in sympathetic nervous system tone (SNS) which, in concert with TH, accelerates energy
expenditure (EE) and increases body temperature3. Numerous studies primarily performed on animal models have investigated the mechanism of TH for activating brown adipose tissue (BAT).
Findings from different TR knockout mice (e.g. TRα, TRβ, TRα1+m) and the use of isoform-selective agonists indicate specific roles for TH-adrenergic interactions in BAT and white adipose
tissue(WAT)4,5,6,7,8,9,10. Early studies in thyroidectomized and hypothyroid rats demonstrated the relevance of BAT specific type II iodothyronine deiodinase Dio2 as a local and systemic
source of 3,3′,5-triiodothyronine (T3) under cold exposure and for the first time provided evidence for T3 in the enhancement of the thermogenic response to sympathetic stimulation11,12.
This suggests that the subsequent heat liberation is actually a result of synergism between norepinephrine (NE) and T3 signaling13. The significance of the central nervous system in TH
activation of BAT was demonstrated in rats in which central administration of T3, via the inhibition of hypothalamic AMP-activated protein kinase (AMPK), induces thermogenic gene expression
in BAT through stimulation of sympathetic nervous system (SNS) activity14. Additionally, SNS stimulation results in activation of β-adrenergic receptors (β-AR) by NE which induces both the
enhanced thermogenic activity of existing brown adipocytes and the recruitment of new cells to BAT depots1. Energy release is accomplished by the activity of UCP1, a BAT-specific
mitochondrial uncoupling protein1. With regards to WAT, a distinct population of UCP1-positive adipocytes arises within subcutaneous and visceral depots of mice in response to cold
exposure15,16 and β3 adrenergic receptor agonist treatment15,17,18. To date it remains a matter of controversy as to whether these cells, entitled “brite” or “beige” adipocytes, are produced
from the conversion of existing white adipocytes and/or the differentiation of WAT resident progenitor cells, leading to a process called “browning”19,20,21. There is preliminary evidence
that suggests a role for TH in the induction of beige adipocytes in WAT. In human multipotent adipose-derived stem cells, 3–10 day treatment with T3 induced UCP1 expression which was most
pronounced during the differentiation phase and was dependent on the presence of thyroid hormone receptor β (TRβ)22. Similarly, treatment of mice with the TRβ specific agonist GC-1 leads to
induction of adaptive thermogenesis in subcutaneous WAT, increased energy expenditure, and restoration of cold tolerance in cold-intolerant mice independent from BAT activity23,24.
Furthermore, in mice centrally administered T3 leads to the recruitment of BAT in inguinal WAT which is accompanied by increased energy expenditure and body temperature. Since the effect was
abolished in UCP1 KO mice, the findings highlight the significance of UCP1 as a central relais modulating the central effects of TH25. Furthermore, a novel circuit for the control of TH
feedback in the brain via the Liver X receptor α and β has recently been published26. LXRαβ−/− mice display increased TH serum level combined with changes in expression of genes associated
with TH synthesis and TH transport as well as prominent UCP1 staining in the subcutaneous adipose depot26. In a positron emission tomography (PET) study of a patient with a history of
thyroid cancer, systemic administration of TH led to BAT activation in periumbilical WAT indicating that browning had occurred as a result of T4 supplementation for 14 days27. Despite the
significance of TH for adaptive thermogenesis, there is currently limited understanding of whether and how different thyroid states, i.e. hypo- and hyperthyroidism, regulate BAT
thermogenesis as well as the thermogenic activity of WAT. Therefore, the present study aimed to determine TH induced effects on (I) BAT activity and (II) WAT browning in a mouse model of
thyroidal dysfunction with a view to incorporate the results into the context of TH-regulated effects on whole-body energy homeostasis. RESULTS BOTH HYPO- AND HYPERTHYROIDISM INDUCE BROWN
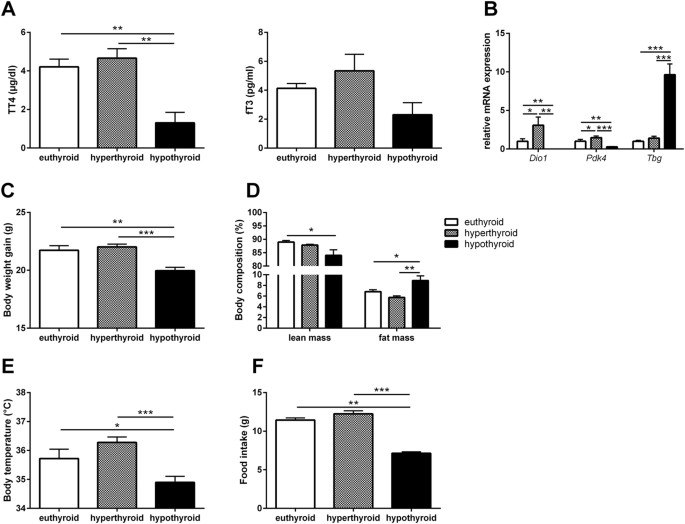
FAT BIOGENESIS Comparison of total T4 and free T3 serum concentrations between eu-, hyper- and hypothyroid mice confirmed the hyperthyroid and hypothyroid state of the animals (Fig. 1A).
This finding is further substantiated by the differential hepatic expression of the TH-responsive genes deiodinase 1 (_Dio1_), thyroxine-binding globulin (_Tbg_) and pyruvate dehydrogenase
kinase 4 (_Pdk4_) (Fig. 1B). The mRNA level of _Dio1_ and _Pdk4_ in liver increased significantly upon T4-treatment and decreased significantly in hypothyroid animals. In addition, induction
of _Tbg_, negatively regulated by TH, confirmed the hypothyroid state (Fig. 1B). Hyperthyroid mice showed slightly enhanced weight gain at the end of treatment while hypothyroid animals
displayed significantly decreased body weights (Fig. 1C). While there was no change in body composition between hyperthyroid and euthyroid animals, hypothyroid mice had significantly reduced
lean mass and concurrently increased body fat (8.9 ± 0.9%) compared with euthyroid (6.8 ± 0.3%) and hyperthyroid mice (5.7 ± 0.3%; Fig. 1D). Core body temperature was significantly
decreased in hypothyroid mice (34.9 ± 0.2 °C) compared with hyperthyroid (36.2 ± 0.2 °C) and euthyroid mice (35.7 ± 0.3 °C); Fig. 1E). Furthermore, food intake decreased in hypothyroid
animals (Fig. 1F). In view of the striking differences in core body temperature and body fat distribution between hyper- and hypothyroid mice, we next asked to which extent WAT depot
contributes to TH induced thermogenesis. Consistent with the finding of increased fat mass, adipocytes from hypothyroid mice were larger in size with increased lipid accumulation in iWAT and
gWAT when compared with hyper- and euthyroid mice (Fig. 2A). In addition we observed moderate increases in thermogenic gene program in the white fat depots of both hypo- and hyperthyroid
mice as compared to euthyroid littermates. Strikingly, _Ucp1_ gene expression was upregulated by about 40-fold in iWAT of hypo- and hyperthyroid mice compared to euthyroid controls.
Moreover, we observed a significant increase in _Ucp1_ mRNA in gWAT, which is considered to have a much lower capacity to induce thermogenic gene programs (Fig. 2B). Of interest, the zinc
finger protein _Zfp423_, which is an essential determinant of preadipocyte commitment28,29, was predominantly upregulated in WAT of hypothyroid mice (Fig. 2B). Histological examination of
the white fat depots revealed the presence of abundant islets with a distinctive multilocular appearance of adipocytes which were positively stained for UCP1 in both, hyper- and hypothyroid
states (Fig. 3A). We then investigated whether the presence of brown fat-like cells has consequences for fatty acid oxidation. As demonstrated in Fig. 3B, the highest uptake of 14C-acetate
was found in both iWAT and gWAT of hyperthyroid mice in comparison with hypo- and euthyroid mice. This was accompanied by a significantly higher level of _Ardb1_ expression in iWAT of
hyperthyroid mice, whereas the expression _Ardb1_ and _Ardb3_ in gWAT of hypo- and euthyroid mice did not change (Fig. 3C). HYPOTHYROIDISM IS ASSOCIATED WITH ENHANCED Β3-ADRENERGIC TONE IN
BAT Given the differences in core body temperature despite the activation of the thermogenic program in WAT at room temperature, we next asked whether this is due to changes in the
thyroid-adrenergic axis induced by thyroidal dysfunction. Immunohistochemical staining of tyrosine hydroxylase, which is the rate-limiting enzyme for catecholamine synthesis30, revealed no
differences in abundance in the BAT of hyperthyroid and hypothyroid mice (data not shown). Furthermore, while we did not observe changes in gene expression of the β-adrenergic receptor 1
(_Ardb1_) in BAT, there was a significant increase in gene expression of _Ardb3_ in BAT of hypo- vs hyperthyroid mice (Fig. 4A). BAT is profusely innervated by sympathetic nerve terminals
with norepinephrine (NE) acting via β-ARs31. Therefore, we next determined the concentrations of circulating NE and epinephrine in the experimental groups. Intriguingly, we found that
concentrations of NE only increased in hypothyroid mice whereas epinephrine increased in both hyperthyroid and hypothyroid mice compared to euthyroid mice (Fig. 4B). The activity of the Dio2
in hypothyroid BAT was 20-fold increased compared to hyperthyroid mice (p < 0.05). In hyperthyroid BAT Dio2 activity was significantly lower than in euthyroid controls (p < 0.01; Fig.
4C). DIFFERENT MORPHOLOGY AND GENE EXPRESSION IN BAT OF HYPO- AND HYPERTHYROID MICE In order to further unravel the discrepancies between hypothermia despite increased adrenergic signalling
and WAT browning in hypothyroid mice we next investigated BAT metabolism. Histological examination of interscapular BAT revealed gross differences in cell morphology. Whereas BAT of
euthyroid mice contained mixed regions of white and brown adipocytes, the BAT of hypothyroid mice contained predominantly adipocytes with unilocular lipid droplets of intermediate size
between WAT and BAT. Hyperthyroid BAT displayed a distinct morphology with a decreased cell size of the mainly multilocular adipocytes (Fig. 5A+B). Gene expression analysis of thermogenic
markers, including _Ucp1_, _Fgf21, Cidea, Dio2 and Elovl3,_ revealed a remarkable collective overexpression in the hypothyroid BAT compared with hyperthyroid mice (Fig. 5C). However, the
high induction of _Ucp1_ mRNA in hypothyroid mice was not reflected on the level of UCP1 protein expression. Furthermore, there was no difference in UCP1 protein expression between
hyperthyroid and euthyroid mice (Fig. 5D). In contrast, in hyperthyroid BAT we found an increased activation of β-adrenergic signaling as demonstrated by higher gene expression of the
hormone-sensitive lipase (_Hsl_) and adipose triglyceride lipase (_Atgl;_ Fig. 5C) together with an increased Ser660-phosphorylation of HSL (Fig. 5D). VOLUME AND ACTIVITY OF BAT IS
DISTINCTIVELY REGULATED IN HYPO- AND HYPERTHYROID MICE Given the magnitude of thermogenic gene expression in BAT of hypothyroid mice together with the increased stimulation of the
TH-adrenergic axis, hypothyroid mice cannot compensate for hypothermia. Therefore we next addressed whether this is due to defects in BAT activity. First, to determine the oxidative capacity
in BAT, we performed _ex-vivo_14C-acetate uptake assay and observed a significant increase in fatty acid oxidation in BAT of hyperthyroid mice (74.0 ± 14.0 dpm/count) as compared to
hypothyroid mice (18.9 ± 1.95 dpm/count; p < 0.01). There was no significant difference between hypo- and euthyroid mice (47.2 ± 14.5 dpm/count; Fig. 6A). Second, we used a combined PET-
and MR-based delineation of active BAT to investigate whether there are differences in BAT formation between the experimental mouse groups. Hyperthyroid mice contained a higher volume of
active iBAT (0.06 ccm ± 0.01 ccm) when compared to euthyroid (0.04 ccm ± 0.01 ccm; p < 0.05) and hypothyroid mice (0.03 ccm ± 0.01 ccm; Fig. 6B). We then asked whether the increase in
amount of BAT also correlates with increased BAT activity. Metabolically active BAT depots with symmetrical uptake of 18F-FDG were observed in all three groups being most marked in the
hyperthyroid group (Fig. 6C). The SUVR of 18F-FDG accumulation in iBAT was significantly decreased in hypothyroid mice vs. euthyroid mice (8.9 ± 1.3 vs. 18.9 ± 3.8; p < 0.05; Fig. 6D).
Furthermore, there was a trend towards an increased 18F-FDG accumulation in hyperthyroid compared to euthyroid and hypothyroid mice mice (21.99 ± 5.0 vs. 18.9 ± 3.8 and 8.9 ± 1.3, p = 0.06,
respectively). Finally, to determine to which extent the MR based iBAT volume is metabolically activated, the respective 18F-FDG avid iBAT with a SUV threshold of >2 was adjusted to the
MR based iBAT volume. Hyperthyroid mice presented with an active iBAT fraction of 94% the highest BAT activity. In contrast, hypothyroid mice showed only in 81% of the interscapular adipose
tissue relevant metabolic activity. In iBAT of euthyroid mice we found that 92% is activated (Fig. 6E). DISCUSSION The present study provides new evidence of how TH affects thermogenesis in
both brown and white adipose depots. For the first time with regard to metabolic activity of BAT, we were able to identify a distinct profile in hyper- and hypothyroid mice at room
temperature. First, regarding the amount of BAT, the _in vivo_18F-FDG PET/MRI-estimated BAT volumes gave first evidence for an increased total amount of BAT in hyperthyroid mice compared to
hypo- and euthyroid animals (Fig. 6B). Second, although hypo- and euthyroid animals contained detectable BAT, hyperthyroid mice displayed an increased metabolic activity as determined by
18F-FDG uptake (Fig. 6C+D). In line with this observation, fatty acid oxidation activity in BAT was increased in hyperthyroid mice as determined by the uptake of 14C-acetate (Fig. 6A). The
opposite metabolic profile was found in hypothyroid animals, which display significantly lower uptake of 18F-FDG and of 14C-acetate in BAT when compared to hyperthyroid mice (Fig. 6A–D). The
observation of active BAT in all three experimental groups emphasizes the significance of thermal conditions for metabolic control irrespective of thyroidal state. As discussed by
Nedergaard _et al_. under sub-thermal conditions (for mice 18–22 °C) BAT derived heat will be used to maintain body temperature32. Paradoxically, the low BAT activity in hypothyroid animals
was associated with a strong increase of thermogenic genes (_Ucp1_, _Dio2_, _Fgf21_, _Cidea_, and _Elovl3_; Fig. 5) and with a significant upregulation of the adrenergic system, as evidenced
by significant increases of _Ardb3_ and NE concentrations in hypothyroid animals (Fig. 4). Although NE _per se_ is not an index of NE release or sympathetic tone, these data suggest an
increase in norepinephrine outflow to the periphery as a compensatory response to maintain body temperature. This finding is principally in agreement with observations in cold exposed
hypothyroid rodents6,33. Interestingly, in iWAT and gWAT of hypothyroid mice we detected features of adipose tissue browning, evidenced by an increased expression of brown specific genes
(_Ucp1_, _Cidea_, and _Elovl3_; Fig. 2) and a multilocular UCP1-positive phenotype (Fig. 3). Collectively, these data suggest a compensatory WAT browning as a response of decreased heat
production due to BAT inactivity in hypothyroid mice. Indeed, decreased BAT thermogenesis in mice lacking bone morphogenetic protein (BMPR1A) is associated with increased circulating NE and
a compensatory browning of iWAT and eWAT in a very similar fashion to the hypothyroid mice in the present study34. The idea of heat compensation is further substantiated by the observation
of enriched expression of the _zinc finger protein 423_ (_Zfp423_) in WAT and BAT selectively in hypothyroid mice (Figs 2+5). Gupta _et al_. demonstrated that _Zfp423_+ endothelial cells
undergo an endothelial-mesenchymal transition into _Zfp423_+_Pparg_+pericytes that serve as a pool of committed pre-adipocytes29. Thus, the increased _Zfp423_ expression in the adipose
tissues of hypothyroid mice likely suggests the formation of beige adipocytes by recruitment and _de-novo_ differentiation of progenitor cells as has been demonstrated by cold exposure or
adrenergic _β__3__-AR_ agonist treatment28. The observation of browning of white adipose tissue was also made in white adipose tissues of hyperthyroid mice, where in particular, established
markers for adipose tissue browning such as _Ucp1, Fgf21_, _Cidea_, and _Elovl3_ were upregulated (Fig. 2). In addition, hyperthyroid mice were characterized by a significant increase in
_Ardb1_ expression in iWAT (Fig. 3C). Recent studies demonstrated that the absence of the _β__1__-AR_ receptor impairs NE-induced _de-novo_ brown adipogenesis in BAT35. Conversely,
_β__1__-AR_ transgenic mice are resistant to diet-induced obesity and display a high abundance of adipocytes expressing Ucp1 in WAT36. With the results gained in the present study we cannot
conclude whether or not central effects of T3 contribute to the observed WAT browning. However, evidences from a recent study by Alvarez-Crespo _et al_. suggest a coordinated action of
central hyperthyroidism on WAT browning. Thus, the authors for the first time demonstrated that central T3 administration in mice induces UCP1 dependent thermogenesis in BAT and browning of
WAT25. Also, similar to the observed increased BAT amount in our cohort of hyperthyroid mice, Alvarez-Crespo reported BAT recruitment upon central T3 infusion even at thermoneutrality25.
Notably, keeping in mind that the hyperthyroid state of our mouse cohort is rather mild it is even more interesting to observe the significant effects on BAT recruitment and WAT browning.
Further work will be necessary to address the extent of centrally induced T3 effects, i.e. by the determination of hypothalamic AMPK and ACC in our model of hyperthyroidism37. Collectively,
our data provide first evidence for differential mechanisms contributing to WAT browning dependent upon altered thyroidal state. This is most apparent in hypothyroid mice that exhibit
increased adipogenesis and _de-novo_ differentiation of brown adipocytes potentially as a compensatory mechanism to hypothermia resulting from BAT inactivity. In hyperthyroid mice, it can be
hypothesized that increased β-adrenergic activation contributes to WAT browning most likely by central effects of TH. However, it has to be emphasized that with the current data we cannot
exclude potential non cell-autonomous contributions from the SNS to adipose tissue browning irrespective of thyroidal state. Thus, in order to rule out that temperature as opposed to
thyroidal state is the underlying reason for WAT browning, further studies should be performed under temperatures where sympathetic stimulation of facultative thermogenesis is depressed i.e.
thermoneutrality (30 °C)32. METHODS MICE Nine week-old female C57BL/6NTac mice were purchased from Taconic Europe (Lille Skensved, Denmark). The mice were housed in pathogen-free facilities
in groups of three at 22 °C with a 12:12-h dark-light cycle (lights on at 06:00 h). Guidelines were approved by the local authorities of the State of Saxony, Germany as recommended by the
responsible local animal ethics review board (Regierungspräsidium Leipzig, TVV04/12, Germany). All experiments with mice were carried out according to the approved guidelines. After one week
of adaptation, induction of hyper- or hypothyroidism started at the age of ten weeks with n = 20 mice per experimental group. Mice were rendered hyperthyroid by application of L-thyroxine
at a dose of 2 μg/ml diluted in drinking water for 4 weeks38. Hypothyroidism was induced by feeding an iodine-free chow diet supplemented with 0.15% propylthiouracil (PTU, catalog TD 97061;
Harlan Teklan, Madison, WI, USA) for 4 weeks. Euthyroid mice fed standard chow diet (Altromin GmbH, Lage, Germany) served as controls. BODY COMPOSITION Body weight was recorded once a week
and after 4 weeks of treatment. Whole body composition (fat mass, lean mass and total body water) was determined in conscious mice by using nuclear magnetic resonance technology with an
EchoMRI700™ instrument (Echo Medical Systems, Houston, TX, USA). Ten animals per experimental group were measured. Rectal body temperature was measured in fed mice between 8:00–9:00 am. Mice
were sacrificed by CO2 inhalation at the age of 14 weeks. 18F-FDG PET/MRI OF BAT ACTIVATION In order to investigate the influence of thyroidal status on BAT activity, small animal
PET/magnetic resonance (MR) imaging studies were performed using a dedicated high-resolution scanner (nanoScan, Mediso Medical Imaging Systems, Hungary). Anaesthetized (induction at 4%,
maintenance at 1.8% isoflurane in 60%/40% oxygen/air) mice were injected intraperitoneally with 14.5 ± 1.3 MBq 18F-FDG followed by a PET/MR scan from 30-60 minute _post-injection_ in list
mode. The data were reconstructed dynamically into 5-min time frames (OSEM, 4 iterations, 6 subsets, MR-based attenuation and scatter correction). A T1-weighted MR image (Gradient Echo
sequence (GRE), TE = 3.2 ms (out phase), TE = 6.4 ms (in phase), TR = 15.0 ms, Flip Angle = 25°; NEX = 2) served for anatomical orientation with individual PET datasets as well as for
threshold based delineation of BAT in the intrascapular region. The 18F-FDG uptake was determined using mean standardized uptake values (SUV) in PET/MR based volumes of interest (VOIs) in
the intrascapular BAT (iBAT) and liver. To rule out an impact of alterations in the physiological 18F-FDG distribution associated with a hyperthyroid or hypothyroid state, 18F-FDG uptake was
accessed semi-quantitatively. Therefore, SUV ratios (SUVR) were calculated to normalize iBAT uptake to unspecific liver uptake39,40. Furthermore, iBAT volume was retrieved by a MR-based
delineation of fatty tissue in the intrascapular region. Active iBAT volume was defined as iBAT with an increased 18F-FDG uptake of SUV > 2. Subsequently, the fraction of active iBAT was
calculated as percentage of active iBAT volume out of total adipose tissue volume delineated by MR in the interscapular region. The SUVR and active iBAT volume between the groups were
compared using 1-way ANOVA, followed by Bonferroni’s multiple comparison test, using Prism 6.0 (GraphPad, San Diego, Ca, USA). Values of p < 0.05 were considered significant.
_EX-VIVO_14C-ACETATE LOADING ASSAY To quantify the oxidative capacity of WAT and BAT in hyper-, hypo-, and euthyroid mice, we performed an _ex-vivo_14C-acetate loading assay that measures
the incorporation of radioactively labeled acetate into triacyl glycerides (TAG) as described elsewhere41. ADIPOCYTE CELL SIZE AND NUMBER To determine cell size distribution and adipocyte
number, 200 μl aliquots of adipocyte suspension were fixed with osmic acid, incubated for 48 h at 37 °C, and counted in a Coulter counter (Multisizer III; Beckman Coulter, Krefeld,
Germany)42. SERUM CONCENTRATIONS OF THYROXINE AND SERUM METABOLITES Serum TT4 and fT3 concentrations were measured using commercial ELISA kits according to the manufacturer’s instructions
(DRG Instruments GmbH, Germany). Serum concentrations of epinephrine and norepinephrine were determined using a commercially available ELISA kit (2-CAT (A-N) Research ELISA; Rocky Mountain
Diagnostics, Inc., Colorado Springs, USA). The immunoassays were performed according to the manufacturer’s instructions. DEIODINASE TYPE 2 ACTIVITY Activity of type 2 deiodinase were
determined in duplicates in BAT homogenates (100 μg protein per 100 μl reaction) as described43,44. WESTERN BLOT ANALYSIS Western Blot analysis was performed on samples of inguinal WAT
(iWAT), gonadal WAT (gWAT) and BAT as described previously45. The following antibodies and dilutions were used: mouse polyclonal anti-Ucp1 antibody (1:500; Merck Millipore; Darmstadt,
Germany), pHSL and HSL(1:1000; Cell Signaling Technology, Danvers, USA). Blots were visualized by enhanced chemiluminescence (Pierce, Milwaukee, USA). QUANTITATIVE REAL-TIME-PCR (QPCR) For
quantification of gene expression qPCR was performed using the LightCycler System LC480 and LightCycler-DNA Master SYBR Green I Kit (Roche, Mannheim, Germany) as described previously46.
Primer sequences are available on request. Gene expression was calculated by the delta-delta CT method using _36B4_ as a reference gene47. Relative gene expression was calculated by setting
the mean of the euthyroid control group to 1 and then calculating each individual value of the three groups of mice studied. IMMUNOHISTOCHEMISTRY Tissues were harvested, fixed in 4%
paraformaldehyde (pH 7.4) for 24 h at 4 °C and subsequently embedded in paraffin. Immunohistochemical analysis was performed on BAT and WAT sections rabbit anti-UCP1 polyclonal antibody
(ab23841; Abcam, Cambridge, UK). Sections were rinsed thoroughly and incubated with labelled polymer HRP anti-rabbit (Dako Envision™+; Dako, Hamburg, Germany) for 1 h. Visualization was
performed with 3,3′-diaminobenzidine. Microscopic examination was performed using an Axio Observer Microscop (Carl Zeiss, Jena, Germany). Images were obtained using ZEN2012 software (Carl
Zeiss, Germany). STATISTICAL ANALYSES Data are shown as mean ± SEM. Statistical significance was determined by Student´s t-test for comparison of two experimental groups or by ANOVA for
comparison of multiple conditions followed by Bonferroni’s multiple comparison test, using Prism 6.0 (GraphPad, San Diego, Ca, USA). _P_ values less or equal to 0.05 were considered to be
significant. ADDITIONAL INFORMATION HOW TO CITE THIS ARTICLE: Weiner, J. _et al_. Thyroid hormone status defines brown adipose tissue activity and browning of white adipose tissues in mice.
_Sci. Rep._ 6, 38124; doi: 10.1038/srep38124 (2016). PUBLISHER'S NOTE: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional
affiliations. REFERENCES * Cannon, B. & Nedergaard, J. Brown Adipose Tissue: Function and Physiological Significance. Physiol. Rev. 84, 277–359 (2004). CAS PubMed Google Scholar *
Silva, J. E. The Thermogenic Effect of Thyroid Hormone and Its Clinical Implications. Ann. Intern. Med. 139, 205–213 (2003). CAS PubMed Google Scholar * Himms-Hagen, J. Role of
thermogenesis in the regulation of energy balance in relation to obesity. Can. J. Physiol. Pharmacol. 67, 394–401 (1989). ADS CAS PubMed Google Scholar * Ying, H., Araki, O., Furuya, F.,
Kato, Y. & Cheng, S.-Y. Impaired adipogenesis caused by a mutated thyroid hormone alpha1 receptor. Mol. Cell. Biol. 27, 2359–2371 (2007). CAS PubMed PubMed Central Google Scholar *
Ribeiro, M. O. et al. Thyroid hormone–sympathetic interaction and adaptive thermogenesis are thyroid hormone receptor isoform--specific. J. Clin. Invest. 108, 97–105 (2001). CAS PubMed
PubMed Central Google Scholar * Christoffolete, M. A. et al. Mice with Targeted Disruption of the Dio2 Gene Have Cold-Induced Overexpression of the Uncoupling Protein 1 Gene but Fail to
Increase Brown Adipose Tissue Lipogenesis and Adaptive Thermogenesis. Diabetes 53, 577–584 (2004). CAS PubMed Google Scholar * Sjögren, M. et al. Hypermetabolism in mice caused by the
central action of an unliganded thyroid hormone receptor α1. EMBO J. 26, 4535–4545 (2007). PubMed PubMed Central Google Scholar * Warner, A. et al. Inappropriate heat dissipation ignites
brown fat thermogenesis in mice with a mutant thyroid hormone receptor α1. Proc. Natl. Acad. Sci. USA 110, 16241–16246 (2013). ADS CAS PubMed Google Scholar * Venditti, P. et al. T3 and
the thyroid hormone beta-receptor agonist GC-1 differentially affect metabolic capacity and oxidative damage in rat tissues. J. Exp. Biol. 212, 986–993 (2009). CAS PubMed Google Scholar *
Venditti, P. et al. The TRbeta-selective agonist, GC-1, stimulates mitochondrial oxidative processes to a lesser extent than triiodothyronine. J. Endocrinol. 205, 279–289 (2010). CAS
PubMed Google Scholar * Silva, J. E. & Larsen, P. R. Adrenergic activation of triiodothyronine production in brown adipose tissue. Nature 305, 712–713 (1983). ADS CAS PubMed Google
Scholar * Silva, J. E. & Larsen, P. R. Potential of brown adipose tissue type II thyroxine 5′-deiodinase as a local and systemic source of triiodothyronine in rats. J. Clin. Invest. 76,
2296–2305 (1985). CAS PubMed PubMed Central Google Scholar * Branco, M., Ribeiro, M., Negrão, N. & Bianco, A. C. 3,5,3′-Triiodothyronine actively stimulates UCP in brown fat under
minimal sympathetic activity. Am. J. Physiol. - Endocrinol. Metab. 276, E179–E187 (1999). CAS Google Scholar * Lopez, M. et al. Hypothalamic AMPK and fatty acid metabolism mediate thyroid
regulation of energy balance. Nat. Med. 16, 1001–1008 (2010). CAS PubMed PubMed Central Google Scholar * Guerra, C., Koza, R. A., Yamashita, H., Walsh, K. & Kozak, L. P. Emergence of
brown adipocytes in white fat in mice is under genetic control. Effects on body weight and adiposity. J. Clin. Invest. 102, 412–420 (1998). CAS PubMed PubMed Central Google Scholar *
Cousin, B. et al. Occurrence of brown adipocytes in rat white adipose tissue: molecular and morphological characterization. J. Cell Sci. 103 (Pt 4), 931–942 (1992). ADS CAS PubMed Google
Scholar * Granneman, J. G., Li, P., Zhu, Z. & Lu, Y. Metabolic and cellular plasticity in white adipose tissue I: effects of beta3-adrenergic receptor activation. Am. J. Physiol.
Endocrinol. Metab. 289, E608–616 (2005). CAS PubMed Google Scholar * Ghorbani, M. & Himms-Hagen, J. Appearance of brown adipocytes in white adipose tissue during CL 316,243-induced
reversal of obesity and diabetes in Zucker fa/fa rats. Int. J. Obes. Relat. Metab. Disord. J. Int. Assoc. Study Obes. 21, 465–475 (1997). CAS Google Scholar * Himms-Hagen, J. et al.
Multilocular fat cells in WAT of CL-316243-treated rats derive directly from white adipocytes. Am. J. Physiol. Cell Physiol. 279, C670–681 (2000). CAS PubMed Google Scholar * Petrovic, N.
et al. Chronic Peroxisome Proliferator-activated Receptor γ (PPARγ) Activation of Epididymally Derived White Adipocyte Cultures Reveals a Population of Thermogenically Competent,
UCP1-containing Adipocytes Molecularly Distinct from Classic Brown Adipocytes. J. Biol. Chem. 285, 7153–7164 (2010). CAS PubMed Google Scholar * Rosenwald, M., Perdikari, A., Rülicke, T.
& Wolfrum, C. Bi-directional interconversion of brite and white adipocytes. Nat. Cell Biol. 15, 659–667 (2013). CAS PubMed Google Scholar * Lee, J.-Y. et al. Triiodothyronine induces
UCP-1 expression and mitochondrial biogenesis in human adipocytes. Am. J. Physiol. Cell Physiol. 302, C463–472 (2012). ADS CAS PubMed Google Scholar * Villicev, C. M. et al. Thyroid
hormone receptor beta-specific agonist GC-1 increases energy expenditure and prevents fat-mass accumulation in rats. J. Endocrinol. 193, 21–29 (2007). CAS PubMed Google Scholar * Lin, J.
Z. et al. Pharmacological Activation of Thyroid Hormone Receptors Elicits a Functional Conversion of White to Brown Fat. Cell Rep. 13, 1528–1537 (2015). CAS PubMed PubMed Central Google
Scholar * Alvarez-Crespo, M. et al. Essential role of UCP1 modulating the central effects of thyroid hormones on energy balance. Mol. Metab. 5, 271–282 (2016). CAS PubMed PubMed Central
Google Scholar * Miao, Y. et al. Liver X receptor β controls thyroid hormone feedback in the brain and regulates browning of subcutaneous white adipose tissue. Proc. Natl. Acad. Sci. 112,
14006–14011 (2015). ADS CAS PubMed Google Scholar * Skarulis, M. C. et al. Thyroid Hormone Induced Brown Adipose Tissue and Amelioration of Diabetes in a Patient with Extreme Insulin
Resistance. J. Clin. Endocrinol. Metab. 95, 256–262 (2010). CAS PubMed Google Scholar * Gupta, R. K. et al. Zfp423 Expression Identifies Committed Preadipocytes and Localizes to Adipose
Endothelial and Perivascular Cells. Cell Metab. 15, 230–239 (2012). CAS PubMed PubMed Central Google Scholar * Gupta, R. K. et al. Transcriptional control of preadipocyte determination
by Zfp423. Nature 464, 619–623 (2010). ADS CAS PubMed PubMed Central Google Scholar * Giordano, A., Frontini, A. & Cinti, S. Adipose organ nerves revealed by immunohistochemistry.
Methods Mol. Biol. Clifton NJ 456, 83–95 (2008). Google Scholar * Youngstrom, T. G. & Bartness, T. J. Catecholaminergic innervation of white adipose tissue in Siberian hamsters. Am. J.
Physiol. 268, R744–751 (1995). CAS PubMed Google Scholar * Nedergaard, J. & Cannon, B. The browning of white adipose tissue: some burning issues. Cell Metab. 20, 396–407 (2014). CAS
PubMed Google Scholar * de Jesus, L. A. et al. The type 2 iodothyronine deiodinase is essential for adaptive thermogenesis in brown adipose tissue. J. Clin. Invest. 108, 1379–1385 (2001).
CAS PubMed PubMed Central Google Scholar * Schulz, T. J. et al. Brown-fat paucity due to impaired BMP signalling induces compensatory browning of white fat. Nature 495, 379–383 (2013).
ADS CAS PubMed PubMed Central Google Scholar * Lee, Y.-H., Petkova, A. P., Konkar, A. A. & Granneman, J. G. Cellular origins of cold-induced brown adipocytes in adult mice. FASEB J.
Off. Publ. Fed. Am. Soc. Exp. Biol. 29, 286–299 (2015). CAS Google Scholar * Soloveva, V., Graves, R. A., Rasenick, M. M., Spiegelman, B. M. & Ross, S. R. Transgenic mice
overexpressing the beta 1-adrenergic receptor in adipose tissue are resistant to obesity. Mol. Endocrinol. Baltim. Md 11, 27–38 (1997). CAS Google Scholar * López, M. et al. Hypothalamic
AMPK and fatty acid metabolism mediate thyroid regulation of energy balance. Nat. Med. 16, 1001–1008 (2010). PubMed PubMed Central Google Scholar * Bianco, A. C. et al. American Thyroid
Association Guide to Investigating Thyroid Hormone Economy and Action in Rodent and Cell Models. Thyroid 24, 88–168 (2014). PubMed PubMed Central Google Scholar * Wu, J., Cohen, P. &
Spiegelman, B. M. Adaptive thermogenesis in adipocytes: Is beige the new brown? Genes Dev. 27, 234–250 (2013). CAS PubMed PubMed Central Google Scholar * Skillen, A., Currie, G. M. &
Wheat, J. M. Thermal control of brown adipose tissue in 18F-FDG PET. J. Nucl. Med. Technol. 40, 99–103 (2012). PubMed Google Scholar * Kern, M. et al. Liver-restricted Repin1 deficiency
improves whole-body insulin sensitivity, alters lipid metabolism, and causes secondary changes in adipose tissue in mice. Diabetes 63, 3295–3309 (2014). CAS PubMed Google Scholar *
Ruschke, K. et al. Repin1 maybe involved in the regulation of cell size and glucose transport in adipocytes. Biochem. Biophys. Res. Commun. 400, 246–251 (2010). CAS PubMed Google Scholar
* Leonard, J. L. & Rosenberg, I. N. Iodothyronine 5′-deiodinase from rat kidney: substrate specificity and the 5′-deiodination of reverse triiodothyronine. Endocrinology 107, 1376–1383
(1980). CAS PubMed Google Scholar * Hoefig, C. S. et al. Biosynthesis of 3-Iodothyronamine From T4 in Murine Intestinal Tissue. Endocrinology 156, 4356–4364 (2015). CAS PubMed Google
Scholar * Krause, K. et al. Dissecting molecular events in thyroid neoplasia provides evidence for distinct evolution of follicular thyroid adenoma and carcinoma. Am. J. Pathol. 179,
3066–3074 (2011). CAS PubMed PubMed Central Google Scholar * Krause, K. et al. TFF3-based candidate gene discrimination of benign and malignant thyroid tumors in a region with borderline
iodine deficiency. J. Clin. Endocrinol. Metab. 93, 1390–1393 (2008). CAS PubMed Google Scholar * Livak, K. J. & Schmittgen, T. D. Analysis of relative gene expression data using
real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods San Diego Calif 25, 402–408 (2001). CAS Google Scholar Download references ACKNOWLEDGEMENTS The authors thank Prof.
Dr. M. Patt, Department of Nuclear Medicine, University of Leipzig, Germany, for the supply of radiotracer and Eva Böge and Viola Döbel, Department of Internal Medicine, University of
Leipzig, Germany, for technical assistance. This work was supported by the DFG SPP1629 (TO 718/2, KR425811-1, KO 922/17-1; Z KO 922/17-1/2) as well as the DFG SFB1052 (C01 and C07), from the
German Diabetes Association and from the DHFD (Diabetes Hilfs- und Forschungsfonds Deutschland). AUTHOR INFORMATION AUTHORS AND AFFILIATIONS * Department of Endocrinology and Nephrology,
University Hospital, Leipzig, Germany Juliane Weiner, Nora Klöting, Claudia Gebhardt, John T. Heiker, Susan Kralisch, Michael Stumvoll, Matthias Blüher, Anke Tönjes & Kerstin Krause *
Department of Neuroradiopharmaceuticals, Helmholtz-Zentrum Dresden-Rossendorf, Institute of Radiopharmaceutical Cancer Research, Research Site Leipzig, Leipzig, Germany Mathias Kranz, Winnie
Deuther-Conrad & Peter Brust * University of Leipzig, IFB Adiposity Diseases, Leipzig, Germany Nora Klöting, Mohammed Hankir, Susan Kralisch, Michael Stumvoll, Matthias Blüher, Osama
Sabri & Swen Hesse * German Center for Diabetes Research (DZD), Leipzig, Germany Anne Kunath * Department of Nuclear Medicine, University Hospital, Leipzig, Germany Karen Steinhoff,
Vilia Zeisig, Osama Sabri & Swen Hesse * Institute of Experimental Endocrinology, Charité University Hospital, Berlin, Germany Eddy Rijntjes & Josef Köhrle Authors * Juliane Weiner
View author publications You can also search for this author inPubMed Google Scholar * Mathias Kranz View author publications You can also search for this author inPubMed Google Scholar *
Nora Klöting View author publications You can also search for this author inPubMed Google Scholar * Anne Kunath View author publications You can also search for this author inPubMed Google
Scholar * Karen Steinhoff View author publications You can also search for this author inPubMed Google Scholar * Eddy Rijntjes View author publications You can also search for this author
inPubMed Google Scholar * Josef Köhrle View author publications You can also search for this author inPubMed Google Scholar * Vilia Zeisig View author publications You can also search for
this author inPubMed Google Scholar * Mohammed Hankir View author publications You can also search for this author inPubMed Google Scholar * Claudia Gebhardt View author publications You can
also search for this author inPubMed Google Scholar * Winnie Deuther-Conrad View author publications You can also search for this author inPubMed Google Scholar * John T. Heiker View author
publications You can also search for this author inPubMed Google Scholar * Susan Kralisch View author publications You can also search for this author inPubMed Google Scholar * Michael
Stumvoll View author publications You can also search for this author inPubMed Google Scholar * Matthias Blüher View author publications You can also search for this author inPubMed Google
Scholar * Osama Sabri View author publications You can also search for this author inPubMed Google Scholar * Swen Hesse View author publications You can also search for this author inPubMed
Google Scholar * Peter Brust View author publications You can also search for this author inPubMed Google Scholar * Anke Tönjes View author publications You can also search for this author
inPubMed Google Scholar * Kerstin Krause View author publications You can also search for this author inPubMed Google Scholar CONTRIBUTIONS J.W. and K.K. designed and performed the
experiments, interpreted and analyzed the data, and wrote the manuscript. N.K. and A.K. performed mouse work. M.K., K.S., V.Z., S.H., W.D.C., P.B., and O.S. performed and analyzed PET/MRI
analysis. E.R. and J.K. determined serum concentrations of thyroid hormones and DIO2 activity assays. C.G., S.K., J.T.H. and M.H. performed experiments and analyzed data. M.B., M.S., and
A.T. interpreted and analyzed the data, and critically edited the manuscript. All authors read, commented and approved the final version of the manuscript. K.K. is responsible for the
integrity of the work as a whole. ETHICS DECLARATIONS COMPETING INTERESTS The authors declare no competing financial interests. RIGHTS AND PERMISSIONS This work is licensed under a Creative
Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in
the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of
this license, visit http://creativecommons.org/licenses/by/4.0/ Reprints and permissions ABOUT THIS ARTICLE CITE THIS ARTICLE Weiner, J., Kranz, M., Klöting, N. _et al._ Thyroid hormone
status defines brown adipose tissue activity and browning of white adipose tissues in mice. _Sci Rep_ 6, 38124 (2016). https://doi.org/10.1038/srep38124 Download citation * Received: 25
February 2016 * Accepted: 07 November 2016 * Published: 12 December 2016 * DOI: https://doi.org/10.1038/srep38124 SHARE THIS ARTICLE Anyone you share the following link with will be able to
read this content: Get shareable link Sorry, a shareable link is not currently available for this article. Copy to clipboard Provided by the Springer Nature SharedIt content-sharing
initiative