Details on the transport of european eel larvae through the strait of gibraltar into the mediterranean sea
- Select a language for the TTS:
- UK English Female
- UK English Male
- US English Female
- US English Male
- Australian Female
- Australian Male
- Language selected: (auto detect) - EN
Play all audios:

ABSTRACT Numbers of European glass eels (_Anguilla anguilla)_ monitored along the Atlantic and Mediterranean coasts of Europe currently serve as the main stock indicator in assessment of
this critically endangered species. Spawning, however, takes place exclusively in the Sargasso Sea, several thousand kilometers away. The beginning of its complex lifecycle is characterized
by a distant and lengthy larval drift, before the young-of-the-year reach the monitoring stations at the European coasts. The oceanic mechanisms regulating dispersal and distribution of
European eel leptocephalus larvae, before they metamorphose into glass eels and colonize future growth habitats, are still poorly understood and data are scarce. Here, we present
oceanographic and leptocephalus catch data from a 24-h station on board of the German Research Vessel Meteor, covering one event cycle of the tide-derived change of hydrographic conditions
in the central part of the Strait of Gibraltar. Results of this study provide detailed insights on how the exchange of water masses between the Atlantic and the Mediterranean Sea may favor
or prevent transport and migration of eel larvae through the Strait, which potentially plays a decisive role in timing and magnitude of larval recruitment events into the entire
Mediterranean region. SIMILAR CONTENT BEING VIEWED BY OTHERS NORTH EQUATORIAL CURRENT AND KUROSHIO VELOCITY VARIATIONS AFFECT BODY LENGTH AND DISTRIBUTION OF THE JAPANESE EEL _ANGUILLA
JAPONICA_ IN TAIWAN AND JAPAN Article Open access 21 February 2022 FIRST DIRECT EVIDENCE OF ADULT EUROPEAN EELS MIGRATING TO THEIR BREEDING PLACE IN THE SARGASSO SEA Article Open access 13
October 2022 NEW CLUES ON THE ATLANTIC EELS SPAWNING BEHAVIOR AND AREA: THE MID-ATLANTIC RIDGE HYPOTHESIS Article Open access 06 October 2020 INTRODUCTION The stock of the panmictic European
eel (_Anguilla anguilla_) exhibited substantial decline in recent decades and is currently rated as critically endangered by the IUCN Red List of Threatened Species1. Recruitment
time-series for glass eels and yellow eels, monitored along the Atlantic, Baltic and Mediterranean coastlines, currently serve as the basis for the evaluation of the status and development
of the stock2. While these data provide crucial insights into recruitment dynamics, they do not capture all potential threats or changes affecting the whole stock, emphasizing the need for a
comprehensive understanding of the complete life cycle including the leptocephalus stage3,4,5. Especially early life history stages such as eggs and larvae are vulnerable to a number of
additional oceanic and environmental factors6,7,8,9,10,11,12. Investigating and surveying changes in oceanic abundance and distribution of larvae can be valuable to define the role of
oceanic factors in the species decline and provide information on the temporal and regional fluctuations in coastal recruitment7,13. Further, knowledge about the transport and dispersal
mechanisms of leptocephalus larvae in the open North-West Atlantic, but also in the coastal areas after their journey from the Sargasso Sea, are key to understand and predict the magnitude
of yearly recruitment in different areas14. Ocean currents play a decisive role in timing as well as trajectory of the drifting leptocephalus larvae, as distinct current patterns have been
identified that either favor a transport to the North Sea or towards the Mediterranean Sea4,11,15,16,17,18. Besides oceanographic features, larval behavior can also be a key element in this
regard since anguillid leptocephali are not seen as fully passive drifters and observations as well as behavioral studies have shown their ability to actively swim horizontally and to rise
and dive in the water column19,20,21. Even though availability of empiric data on the vertical distribution of leptocephalus larvae in the water column is not extensive, current
understanding is that larger, later stage anguillid leptocephali conduct diurnal vertical migrations (DVM) and are usually found in deeper water layers during daytime but in shallower,
surface-near layers during the night13,20,22,23. Previous studies described the preferred depth during nighttime to range between 50 and 70 m, while the extent of vertical distribution
during daylight seems to increase with body length24. Tesch23 reported large leptocephali close to the European shelf to reside mainly between 300 and 600 m during daytime. Previous studies
have shown that this distinct behavior also mirrors in the catchability of the larvae in certain depth ranges, which is the reason why the sampling strategies and targeted depth strata for
specimens of this life history stage usually differ during night and day22,25. The vertical position of the larvae during different times of day and night thus potentially affects their
oceanic distribution, as direction and velocity of currents often vary depending on the depth stratum. Not only due to the complexity of larval oceanic dispersal and the wide natural
distribution, which increase the efforts to develop robust monitoring strategies, there remains a paucity of data on recruitment and standing stock in certain parts of the eel’s range. Glass
eel monitoring at selected sites in the Mediterranean and close to the Strait (e.g. at the Guadalquivir River, located on the Spanish Atlantic side) are considered crucial data points to
describe recruitment trends in the Mediterranean Sea26. Still, the Mediterranean Sea is particularly considered inadequately covered by recruitment time-series, even though the area is
thought to be an important habitat for the species. Until today, it is neither entirely clear in what magnitude its production contributes to the spawning stock biomass, nor precisely how
high the natural recruitment is in this area2,13,26,27,28. The Mediterranean Sea and the Atlantic Ocean are connected by the Strait of Gibraltar, a narrow (∼13 km at its narrowest point) and
rather shallow (∼280 m at the shallowest point, the Camarinal Sill) bottleneck, which young eels inevitably need to pass in order to colonize any coastal area or river system of the
Mediterranean or Black Sea. This, of course, is also true for adult silver eels emigrating out of the Mediterranean, in order to successfully conduct their spawning migration24,27,29.
Similar to other gateways between different ocean basins, water exchange, current and tidal systems in the Strait of Gibraltar are complex and regulated by a variety of factors24.
Semi-enclosed basins, such as the Mediterranean, typically display differences in several physical characteristics from the open Atlantic Ocean29. The water balance of the Baltic Sea for
instance, is controlled by outflow of a freshwater and precipitation-fed brackish water column within the upper layer30, while inflow is characterized by high saline water from the Kattegat
in deeper strata31. In contrast, the Mediterranean Sea is considered a concentration basin, which is characterized by water loss due to evaporation exceeding water gain from precipitation
and river runoff. Adding to this, there is a negative net heat budget of the basin, causing an anti-estuarine vertical thermohaline circulation, as deeper, more saline water exits the
Mediterranean, while surface waters from the Atlantic enter the Mediterranean32,33. Additionally, these density-driven currents are superimposed by tidal currents that can exceed two meters
per second and thus strengthen or slow down the eastwards-directed (i.e., towards the east sector, including east, east-northeast, and east-southeast directions) surface inflow, depending on
the phase of the tide. The tidal energetics of the Strait of Gibraltar are complicated and have been intensely studied33,34,35,36,37. However, even though notable scientific effort in
collecting data for this region was made, for a long time, the tidal regime of the Strait was considered still not entirely understood by some authors38. The current flow is assumed to
interact with the Camarinal Sill (shallow part of the Strait) causing surface wave patterns, that are detectable by satellites35,39. The fluctuating conditions and temporarily strong
currents in the Strait in connection with the limited mobility of the few centimeters long leptocephali suggest that recruitment and transport from the Atlantic into the Mediterranean must
be linked to specific conditions, as was already supposed by Tesch et al. in 198640. The significance of the area for the distribution of eel larvae was also highlighted in a dedicated
paragraph in McCleave et al.24, who hypothesized that the tidal conditions in the Strait make a passive transport of relatively shallow residing leptocephali during spring tide not only
possible, but likely. In more detail, and considering the behavioral characteristics of eel larvae (DVM) as well as the oceanographic conditions in the Strait, it was hypothesized that
optimal conditions for the larval transport into the Mediterranean Sea would only be given, when strong eastward surface currents occur at nighttime after sunset, when the eel larvae ascend
to the upper part of the water column. The here presented study builds on this work by providing oceanographic and larval catch data during a 24-h station to complement the hypothesis with
concrete data. The aim of this study was therefore to gain better insight into the transport of anguillid leptocephali at a highly dynamic bottleneck during their transoceanic larval
migration. Results are based on periodical plankton sampling with an Isaacs-Kidd Midwater Trawl (IKMT) in the central part of the Strait of Gibraltar. During a full tidal cycle, we monitored
velocity in the water column using acoustic doppler current profiler (ADCP), additional hydrographic parameters using a conductivity, temperature and depth rosette (CTD) as well as surface
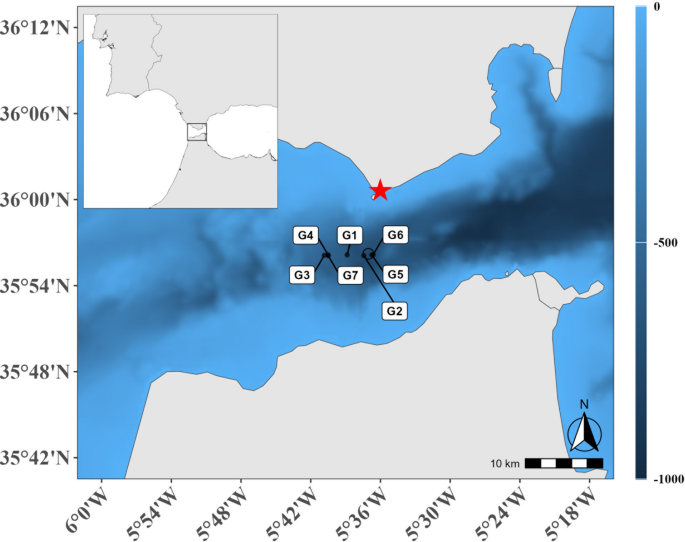
current velocity and direction derived from high-frequency radar (HFR). RESULTS Seven deployments of the IKMT during 24 h in the central part of the Strait of Gibraltar (Fig. 1) revealed
clear differences in leptocephalus catch quantities. While zero larvae were caught during the two daytime and one evening hauls in depths from 0 down to 200 m, a total of 31 leptocephali
including 17 _Anguilla anguilla_ (total length (TL): 54–65 mm), 12 _Conger conger_ (TL: 50–71 mm), 1 _Eurypharynx pelecanoides_ (TL: 16 mm) and 1 _Gnathophis_ sp. (TL: NA) specimen were
caught in the upper 100 m during the four nighttime hauls. A total of 13 _A. anguilla_ and 4 other leptocephali were caught during one (G5) of the deployments on November 21st beginning at
01:00 UTC. This haul produced the highest catch per unit effort (CPUE) of 9.51 caught larvae per hour, as well as the highest calculated larval density (3.31*10−4 individuals / m3) in this
study and the entire survey41 . WEATHER AND OCEANIC CONDITIONS Wind direction and speed (Table 1) during the observed time frame were moderate and tidal current velocities can be considered
common for the Strait at this time of the year. The tides on November 20th and 21st, 2022 occurred during a waning moon, meaning that sampling did not take place during extreme tidal ranges
like those seen during spring tides (Table 2). Higher windspeeds and/or spring tide conditions would have had an increasing effect on the currents in the Strait. WATER STRATIFICATION IN
OBSERVED TIMEFRAME CTD data (Fig. 2) confirmed a stratified water column in the converging area in the central part of the Strait of Gibraltar, which corresponded well with the known
separation of water masses from the Atlantic and the Mediterranean Sea. While the surface layer was characterized by a 70–100 m deep lens of warmer, less saline and less dense water, the
deeper layers of the water column were characterized by more saline, colder and thus denser water masses. In addition to these characteristics, higher oxygen levels in the surface layer
indicate a stronger mixing in this depth stratum. The CTD profiles at 00:04, 06:00 and 13:16 UTC (Fig. 2) reflect the tidal influences on the water stratification (Fig. 3). The profile at
00:04 (purple) was recorded shortly after high tide and showed lower temperatures and higher salinity between 37 and 250 m depth due to the westward transport of Mediterranean water. In
contrast, the profile at 06:00 (orange) was recorded after low tide, with the influence of eastward flowing Atlantic water leading to higher temperatures and lower salinity in the same depth
range. The profile at 13:16 (black), taken after the second high tide, was consistent with the observed westward transport below 300 m. These fluctuations, which are supported by ADCP
measurements (Fig. 3), illustrate the influence of tidal cycles on the distribution of water masses. WATER CURRENT VELOCITY AND DIRECTION MEASURED BY ACOUSTIC DOPPLER CURRENT PROFILER (ADCP)
For technical reasons, the shipboard ADCP velocity measurements started with haul G2 in the afternoon of November 20th and (in correspondence with CTD data) also showed a stratified current
structure in the water column. Strong variation was present in the eastward velocity component (Fig. 3a,c) that flowed from the open Atlantic towards the Mediterranean Basin and rather
negligible variation in the northward velocity components (Fig. 3b,d). For the eastward component, current velocities generally differed during the observed timeframe depending on depth,
with highest velocities in the upper 100 m, reaching a maximum of almost 200 cm s−1 due to strong inflow from the Atlantic into the Mediterranean (Fig. 3a). High velocities of more than 120
cm s−1 were observed at the end of the first monitored outgoing tidal cycle (20/11/2022, approx. 16:20 UTC) until shortly after the first low tide (20/11/2022, approx. 16:50 UTC), as well as
between the first high tide until after the second low tide of the observed timeframe (21/11/2023, approx. 00:15–08:00 UTC). This is highlighted in Fig. 3c for the exemplary depths 37 m
(black) and 101 m (violet), starting around 00:00 UTC on November 21st until after low tide, approx. 08:00 UTC on November 21st, which was shortly after the last haul (G7). Deeper water
strata, exemplified here by the depth of 197 m (orange), show a negative eastward movement (i.e., a westward movement) during most of the tidal cycle, and neutral or only very slight
eastward movement occurring between high tide and low tide (Fig. 3a,c). ADCP vertical profiles of water velocity magnitude and direction during the timing of the most successful haul (G5)
depict the magnitude and direction of the inflow event in eastward direction (Figs. 3, 4c). Water profile plots of current speed and direction at three exemplary times (beginning, mid-haul,
end of haul) of hauls with caught anguillid larvae (G3 (n = 2), G4 (n = 2) and G5 (n = 13)) further depict the respective prevailing current regimes in different depths (Fig. 4a–c). While
current direction and speed during the first haul with caught larvae in the upper water layer (37–100 m) in the beginning and mid haul was directed eastward to southward at a moderate
velocity ranging from 25 to 50 cm s−1 (Fig. 4a), the water turned directions amidst the haul completely westwards through the entire water column from surface down to 600 m (Fig. 4a). The
second haul with caught larvae (Fig. 4b) displayed a westward direction in the very upper water layer and a south-western trajectory in the beginning and mid haul. Current speed was slow to
moderate, ranging from approx. 10–76 cm s−1, with peaks around 100 cm s−1 between 100 and 165 m of depth. The highest observed current velocities were seen in G5, the haul with the highest
count of collected anguillid leptocephali (Fig. 4c). During the three exemplary times of this haul, current velocity reached 175 cm s−1 in the upper 100 m of the water column in an eastward
direction. Current direction in greater depths during this haul turned from north-eastwards to eastwards at low to moderate speed ranging from 0 to 40 cm s−1 (Fig. 4c). ATLANTIC JET SEA
SURFACE CURRENT DERIVED FROM HIGH-FREQUENCY RADAR Data derived from high-frequency radar monitoring sea surface currents in the Strait of Gibraltar show a strong inflow event from November
20th, 2022 at 23:00 UTC until November 21st, 2022 at 08:00 UTC (Fig. 5). High eastward velocities (yellow & red) characterized the Atlantic jet current, which moved surface water masses
eastwards through the Strait of Gibraltar from the Atlantic into the Mediterranean (Fig. 5c,d). DISCUSSION This study is the first to provide catch data and environmental data as connected
evidence of how eel larvae are transported from the Atlantic to the Mediterranean Sea. We provide further support for the hypothesis by McCleave et al.24 that optimal conditions for larval
transport into the Mediterranean Sea arise when strong eastward surface currents occur at night, coinciding with the ascent of eel larvae into the upper, shallower layers of the water column
as part of their diel vertical migration behavior. Utilizing this ‘perfect wave’ of strong eastward currents appears to facilitate the most effective transport of larvae. However, it is
important to note that the necessary conditions are not uniform throughout the year. In winter, southern Spain experiences longer periods of darkness (nearly 14 h) compared to summer. This
extended darkness aligns with tidal cycles, providing more potential windows for larval eel transport than in summer. The exact timing of larval transport within the water column, and
whether it occurs right after sunset or a later at night, may affect their interaction with tidal conditions. If larvae migrate upwards shortly after sunset and return to deeper waters at
sunrise, winter conditions with longer nights could offer more favorable conditions for transport compared to summer, especially during spring tides or strong westerly winds. Miller et al.4
published a comprehensive review on the distribution, timing and size of _A. anguilla_ leptocephali, documenting that late stage leptocephali have been collected almost throughout the entire
year around the Strait of Gibraltar. It is unclear if the abundance of leptocephalus larvae in this area greatly changes throughout the year, or if there is a potential peak of arrival at a
specific timeframe. But while it seems reasonable to assume that there might be a temporal connection to a time before the peak arrival of glass eels in estuaries around the Strait and in
the Mediterranean, it remains unclear if and to what extent larval abundances in this area change throughout the year. Relating the ADCP and HFR data with the IKMT catches of this study,
larvae were only collected at nighttime and highest leptocephalus abundance (G5) was recorded during the formation of an eastward directed wave of Atlantic water towards the Mediterranean
(Figs. 2, 4c). The other two hauls (G3 and G4) containing _Anguilla_ larvae were conducted during non or only slowly westward flowing incoming tide, as can be seen in Fig. 4. These larvae
were possibly in the surface strata of a water mass that has not been entirely pushed through the Strait but instead was (at least in parts) pulled back to the western Atlantic part of the
Strait when the tides changed. The outflow of the heavier, more saline water out of the Mediterranean Sea through the Strait in westward direction possibly constitutes a natural barrier for
leptocephalus larvae in front of the Strait. Miller20 underlined how poorly documented the swimming capabilities of leptocephalus larvae were. Even though it cannot be ruled out that
anguillid eel larvae are capable of single-direction swimming to increase recruitment success19, their capability of actively swimming long distances or against strong currents has been
subject to scientific discussion and speculation in the past4,15,24,40. Only few publications provide empiric data on the swimming capacity of anguillid larvae: Artificially hatched 30–60 mm
long larvae of the Japanese eel (_Anguilla japonica_) reached swimming speeds of 3.6 + /- 2.7 cm s−1 horizontally and 2.8 + /- 1.1 cm s−1 vertically in captivity, which corresponds roughly
to one body length s−121. Interestingly, this constitutes a swimming speed that has shown the best hypothetical recruitment success in models, compared to inactive drifting42. Swimming
speeds of late stage or metamorphosing _Conger oceanicus_ leptocephali and _A. rostrata_ glass eels, measured in a laboratory swimming channel, were faster: ranging from 12.0 to 26.8 cm s−1
and 4.1 to 25.0 cm s−1, respectively43. During the 24 h observation period, current velocities in deeper, westward flowing water layers ranged from approx. 50–100 cm s−1, except during the
tidal jet surface inflow associated with Atlantic high tides. These current velocities, compared to the known swimming speeds of larvae, suggest that active swimming of anguillid
leptocephali against the outflowing currents of the Mediterranean water during daytime over a longer time span and distance must be deemed unrealistic. Tesch et al.40 were apparently the
first to address this question after a larval survey in the area and described a stowing or bottleneck effect in the funnel between Europe and Africa, just before the Strait of Gibraltar.
The authors also concluded that a swimming speed of at least 20 cm s−1 was required to effectively compensate westwards outflowing currents from the Mediterranean. Our data support this, as
the observed tidal dynamics, which are characterized by fast eastward-flowing surface water strata and stationary or slowly westwards-moving deeper layers could explain accumulation effects
of leptocephali west of the Strait. These accumulations likely persist until the next eastward tidal pulse transports the larvae during their shallow nighttime phase of the DVM. Current
direction and velocity in the Strait of Gibraltar are greatly influenced by the Atlantic tidal system, resulting in a constant move of oceanic water from the Atlantic into the Mediterranean
and back. Our recordings during the 24 h station in the Strait are consistent with a model that describes the exchange of water masses between the oceanic bodies as tidal oscillation waves
with movement patterns in different directions, yet mainly following an eastward trajectory44. Izquierdo et al.45 considered tides to be by far the most energetic process in the Strait, that
present a semidiurnal character, though diurnal current velocities were not negligible. These tides are essential for the water movement in the Strait all year round, so although there was
not much wind on this occasion, the observed conditions during the 24 h station can be regarded as representative of average conditions in the Strait of Gibraltar. While most of the time
heavier, more saline water pushes out in deeper layers westwards of the Mediterranean Sea into the Atlantic, high tides and rising water levels in the Atlantic facilitate intensification of
eastward surface currents, pushing into the Strait (e.g. as can be seen in the eastward directed wave in Figs. 2c, 4a–d) with velocities of close to 200 cm s−1 (e.g. 7.2 km h−1 or > 20 km
in three hours tidal phase) and more in the upper 50–100 m of the water column. As during these conditions, also the deeper water layers move eastwards (Fig. 4c), this (albeit considerably
slower) could enable recruitment into the Mediterranean also in greater depths during daytime, when the larvae still linger in deeper waters. Given that the European eel is a panmictic
species, a selective and directed recruitment into the Mediterranean Sea seems unlikely. Yet, the mechanisms, timing and triggers that initiate leptocephalus larvae to metamorphose into
glass eels and colonize continental habitats remain poorly understood. While for glass eels, it was shown that a geomagnetic sense could guide them to use tidal cycles and orientate towards
the coastal shores to colonize coastal and freshwater habitats46, it is unknown whether eel leptocephali have similar capabilities to guide their transoceanic migration47,48. Regardless of
this, larval as well as metamorphosing eels exhibit negative phototaxis, as demonstrated for captive reared Japanese eel larvae, for which overhead light caused clear downward movement in
experimental works21. In the same artificially spawned larvae, Yamada et al.21 observed that swimming speeds during horizontal movement and rising behavior were significantly higher than
during diving behavior and suggested that this may help to regulate distribution during inshore migration of young arriving recruits. Tidal waves pushing surface water through the Strait
into the Mediterranean Sea during nighttime could therefore be regarded as a transport pump that will directly affect dispersal of eel larvae and other organisms conducting nocturnal
vertical migrations into the surface strata of the water column. Macias et al.34, who studied the influence of internal waves on the composition and abundance of zooplankton in the Strait,
already highlighted the need to understand and discuss the physical components and processes when addressing biological patterns in regions with intense hydrodynamic regimes. Despite some
limitations due to the confined timeframe of this study, the obtained data and observations are consistent with expectations and the hypotheses presented herewith. Further research covering
longer timeframes ideally throughout other times of the year together with depth-stratified sampling would help to test the here presented hypotheses. Future studies should include
consideration of peak arrival of recruits in the area, environmental factors such as weather conditions, different tidal phases and changing environmental conditions caused by climate
change. CONCLUSIONS This study describes how the interaction between diurnal vertical migration behavior of leptocephalus larvae and the tidal conditions in the Strait of Gibraltar likely
enables the recruitment of European eel larvae into the Mediterranean Sea. The presented data suggest that a significant eastward surface current in combination with the ascent of
leptocephali into the upper water layers at nighttime effectively transports larvae from the Atlantic Ocean to the Mediterranean Sea via the Strait of Gibraltar. Consequently, the here
presented data contribute to our knowledge about the hydrographic factors determining the magnitude and timing of early life stage recruitment events into the Mediterranean and could help
develop convenient targeted and effective recruitment monitoring strategies for this area, complementary to glass eel recruitment time-series. METHODS The collection of oceanic data, current
parameters, as well as the sampling of leptocephalus larvae, was conducted in November 2022 during one leg of the 185th cruise (M185) aboard the German Research Vessel METEOR. LEPTOCEPHALUS
CATCH DATA All leptocephali in this study were collected using a 1550 µm mesh-size Isaacs-Kidd Midwater Trawl (IKMT) with a ~ 7 m2 mouth opening and a length of 12 m (HYDRO-BIOS Apparatebau
GmbH, Altenholz, Germany). Seven hauls were conducted during a 24-h station between the 20th and 21st of November 2022 in the middle of the Strait of Gibraltar, located roughly at 35°56 N
and 5°37 W (Fig. 1; Table 1). The ships speed over ground was adapted to current speed in order to facilitate the gear’s ideal and comparable trawling speed of 2.5 knots in water. As a
result, starting and ending points but also the timing of vertical positions in each haul varied (See Fig. 1 & S1 for details). Filtered water volumes were assessed with flowmeters
(2030R, General Oceanics, Miami, USA). As stratified sampling was not possible during this cruise, the IKMT was towed in three double oblique hauls between surface and 200 m of depth during
daytime and early evenings and in four quadruple oblique hauls between surface and 100 m of depth during nighttime and early morning, to target only surface-near leptocephali after or during
their diel vertical migration (See Table 1 & S1 for details). This was done to be able to appoint the caught larvae to the upper water strata. Fishing at deeper strata would have
potentially captured deeper larvae and thus may have led to misinterpretation of larval vertical distribution. Collected larvae were immediately sorted from the catch, morphologically
identified, measured, tissue-sampled for genetic species determination and then stored at −20 ℃ for further investigations. All leptocephalus larvae sampled at this station were genetically
confirmed by DNA barcoding. GENETIC SPECIES CONFIRMATION Tissue samples from each leptocephalus larva were separately preserved in ethanol (96% abs.) on board for subsequent genetic
analysis. After the survey, DNA was extracted using Chelex10049 and stored at 4 °C for several weeks or at −80 °C for long-term storage. The verification of morphologically identified
Anguillid species was carried out by a Real-Time PCR-based assay, detecting _A. anguilla_ and _A. rostrata_in comparison to reference samples. All remaining leptocephalus larvae were
morphologically identified, with their species identity confirmed through Nucleotide BLAST50 analysis of the mitochondrial DNA barcoding marker gene Cytochrome c oxidase I (COI), which was
amplified by PCR and sequenced using Sanger sequencing (Service lab: StarSEQ GmbH, Mainz, Germany). OCEANOGRAPHIC DATA As a standard procedure, prior to every deployment of the IKMT during
the 24-h station, vertical observations of temperature (℃), salinity (PSU), water density (kg m−3) and oxygen (mL L−1) were collected by using a SBE 911 conductivity temperature-depth
measure (CTD) (Sea-Bird Electronic, Bellevue, WA, USA). All CTD casts were conducted from sea surface to 600 m, respectively. ADCP DATA Ocean current velocities (from approx. 37 m to the sea
floor, depending on the region and sea state) were collected along the cruise track by a vessel-mounted Teledyne RD Instruments 38 kHz Ocean Surveyor ADCP (Acoustic Doppler Current
Profiler). The shipboard ADCP provides estimates of the horizontal velocity components as a function of depth, based on the Doppler effect of sound wave reflections against small particles
carried by ocean currents. The transducer was located at 5 m below the water line, mounted in the hull of the ship. The instrument was operated in narrowband mode with 16 m bins and a
blanking distance of 16 m, while 100 bins were recorded using a pulse of 2.89 s. The measurements obtained were averaged over 2 min. For more details, see Hanel et al.51. The maximum depth
reached in the Strait of Gibraltar was approx. 626 m. HIGH-FREQUENCY RADAR High-frequency radar (HFR) data was derived from in situ instruments operated by Puertos del Estado in the Strait
of Gibraltar. The HFR system deployed in Strait of Gibraltar consists of four sites equipped with a CODAR-Seasonde model. The sites, owned and operated by Puertos del Estado, started data
collection in May 2011. The system works at a central frequency of 26.8 MHz, providing quality-controlled observations representative of current velocities in the upper 0.5 m of the water
column52. The maximum horizontal range is set to 40 km, and the nominal range and angular resolutions are 1 km and 5°, respectively. This land-based network has proved to be useful in the
characterization of the Atlantic jet53. For further details about the fundamentals and applications in the Mediterranean Sea of this consolidated remote-sensing technology, see Lorente et
al.54. DATA AVAILABILITY The original datasets generated and analyzed during the current study will be made publicly available in the Figshare repository, accessible via
[https://doi.org/10.6084/m9.figshare.c.7583429.v1], upon publication. All relevant data are available from the corresponding author on reasonable request. REFERENCES * Pike, C., Crook, V.
& Gollock, M. _Anguilla anguilla_ (Europe assessment). _The IUCN Red List of Threatened Species_ 2023: e.T60344A216177498 (2023). * ICES. Report of the Joint EIFAAC/ICES/GFCM Working
Group on Eels (WGEEL). _ICES Sci. Rep._ 5, 98 (2023). Google Scholar * Schmidt, J. The breeding places of the eel. _Philos. Trans. R. Soc. Lond. B_ 211, 179–208 (1922). ADS MATH Google
Scholar * Miller, M. J. et al. A century of research on the larval distributions of the Atlantic eels: A re-examination of the data. _Biol. Rev._ 90, 1035–1064 (2015). Article PubMed
Google Scholar * Miller, M. J. et al. Spawning by the European eel across 2000 km of the Sargasso Sea. _Biol. Lett._ 15, 5 (2019). Article MATH Google Scholar * Belpaire, C., Hodson, P.,
Pierron, F. & Freese, M. Impact of chemical pollution on Atlantic eels: Facts, research needs and implications for management. _Curr. Opin. Environ. Sci. Health_ 11, 26–36 (2019).
Article MATH Google Scholar * Bonhommeau, S., Chassot, E. & Rivot, E. Fluctuations in European eel (_Anguilla anguilla_) recruitment resulting from environmental changes in the
Sargasso Sea. _Fish. Oceanogr._ 17, 32–44 (2008). Article Google Scholar * Bonhommeau, S. et al. Impact of climate on eel populations of the Northern Hemisphere. _Mar. Ecol. Prog. Ser._
373, 71–80 (2008). Article ADS MATH Google Scholar * Freese, M. et al. Maternal transfer of dioxin-like compounds in artificially matured European eels. _Environ. Pollut._ 227, 348–356
(2017). Article CAS PubMed MATH Google Scholar * Hanel, R. et al. Low larval abundance in the Sargasso Sea: New evidence about reduced recruitment of the Atlantic eels.
_Naturwissenschaften_ 101, 1041–1054 (2014). Article ADS CAS PubMed MATH Google Scholar * Miller, M. J., Feunteun, E. & Tsukamoto, K. Did a “perfect storm” of oceanic changes and
continental anthropogenic impacts cause Northern Hemisphere anguillid recruitment reductions?. _ICES J. Mar. Sci._ 73, 43–56 (2016). Article Google Scholar * Westerberg, H. et al. Larval
abundance across the European eel spawning area: An analysis of recent and historic data. _Fish Fish._ 19, 324–341 (2018). Article MATH Google Scholar * Westerberg, H. et al. Modeling the
drift of European (_Anguilla anguilla_) and American (_Anguilla rostrata_) eel larvae during the year of spawning. _Can. J. Fish. Aquat. Sci._ 75, 224–234 (2018). Article MATH Google
Scholar * Correia, M. J. et al. The decline in recruitment of the European eel: New insights from a 40-year-long time-series in the Minho estuary (Portugal). _ICES J. Mar. Sci._ 75,
1975–1983 (2018). Article MATH Google Scholar * Bonhommeau, S. et al. The duration of migration of Atlantic _Anguilla_ larvae. _Fish Fish._ 11, 289–306 (2010). Article MATH Google
Scholar * Castonguay, M. et al. Is there a role of ocean environment in American and European eel decline?. _Fish. Oceanogr._ 3, 197–203 (1994). Article MATH Google Scholar * Knights, B.
A review of the possible impacts of long-term oceanic and climate changes and fishing mortality on recruitment of anguillid eels of the Northern Hemisphere. _Sci. Total Environ._ 310,
237–244 (2003). Article ADS CAS PubMed MATH Google Scholar * Munk, P., Mariani, P., Buongiorno Nardelli, B. & Bendtsen, J. Mesoscale driven dispersion of early life stages of
European eel. _Front. Mar. Sci._ 10, 1163125 (2023). Article Google Scholar * Chang, Y.-L.K. et al. Effect of larval swimming in the western North Pacific subtropical gyre on the
recruitment success of the Japanese eel. _PLoS ONE_ 13, e0208704 (2018). Article CAS PubMed PubMed Central Google Scholar * Miller, M. J. Ecology of anguilliform leptocephali:
Remarkable transparent fish larvae of the ocean surface layer. _Aqua BioSci. Monogr._ 2, 1–94 (2009). Article MATH Google Scholar * Yamada, Y., Okamura, A., Mikawa, N. & Utoh, T.
Ontogenetic changes in phototactic behavior during metamorphosis of artificially reared Japanese eel (_Anguilla japonica_) larvae. _Mar. Ecol. Prog. Ser._ 379, 241–251 (2009). Article ADS
Google Scholar * Castonguay, M. & McCleave, J. D. Vertical distributions, diel and ontogenetic vertical migrations, and net avoidance of leptocephali of _Anguilla_ and other common
species in the Sargasso Sea. _J. Plankton Res._ 9, 195–214 (1987). Article Google Scholar * Tesch, F.-W. Occurrence of eel _Anguilla anguilla_ larvae west of the European continental
shelf, 1971–1977. _Environ. Biol. Fish._ 5, 185–190 (1980). Article MATH Google Scholar * McCleave, J. et al. Do leptocephali of the European eel swim to reach continental waters? Status
of the question. _J. Mar. Biol. Assoc. U. K._ 78, 285–306 (1998). Article Google Scholar * Schoth, M. & Tesch, F. W. The vertical distribution of small 0-group _Anguilla_ larvae in the
Sargasso Sea with reference to other anguilliform leptocephali. _Meeresforschung_ 30, 188–195 (1984). MATH Google Scholar * Ciccotti, E. & Morello, E. B. European eel in the
Mediterranean Sea: Outcomes of the GFCM research programme. _FAO Stud. Rev._ No. 103 (forthcoming). * Capoccioni, F. et al. The potential reproductive contribution of Mediterranean migrating
eels to the _Anguilla anguilla_ stock. _Sci. Rep._ 4, 7188 (2014). Article CAS PubMed PubMed Central MATH Google Scholar * Hanel, R. C. et al. Environmental, social and economic
sustainability of European Eel management. _Eur. Parliam. Pol. Dep. Struct. Cohes. Pol._ (2019). * Righton, D. et al. Empirical observations of the spawning migration of European eels: The
long and dangerous road to the Sargasso Sea. _Sci. Adv._ 2(10), e1501694. https://doi.org/10.1126/sciadv.1501694 (2016). Article ADS PubMed PubMed Central Google Scholar * Schinke, H.
& Matthäus, W. On the causes of major Baltic inflows—An analysis of long time series. _Cont. Shelf Res._ 18, 67–97 (1998). Article ADS MATH Google Scholar * Hinrichsen, H.-H. et al.
Sightings of extraordinary marine species in the SW Baltic Sea linked to saline water inflows. _J. Sea Res._ 181, 102175 (2022). Article MATH Google Scholar * Bryden, H. L. & Kinder,
T. H. Steady two-layer exchange through the Strait of Gibraltar. _Deep Sea Res._ 38, 445–463 (1991). Article ADS MATH Google Scholar * Bryden, H. L., Candela, J. & Kinder, T. H.
Exchange through the Strait of Gibraltar. _Prog. Oceanogr._ 33, 201–248 (1994). Article ADS MATH Google Scholar * Macías, D. et al. Chlorophyll maxima and water mass interfaces: Tidally
induced dynamics in the Strait of Gibraltar. _Deep Sea Res._ I(55), 832–846 (2008). Article MATH Google Scholar * Sánchez-Román, A., Criado-Aldeanueva, F., García-Lafuente, J. &
Garrido, J. S. Vertical structure of tidal currents over Espartel and Camarinal sills, Strait of Gibraltar. _J. Mar. Syst._ 74, 120–133 (2008). Article MATH Google Scholar * Vázquez, A.,
Bruno, M., Izquierdo, A., Macías, D. & Ruiz-Cañavate, A. Meteorologically forced subinertial flows and internal wave generation at the main sill of the Strait of Gibraltar. _Deep Sea
Res._ I(55), 1277–1283 (2008). Article Google Scholar * Macías, D. et al. Physical control on zooplankton distribution pattern at the Strait of Gibraltar during an episode of internal wave
generation. _Mar. Ecol. Prog. Ser._ 408, 79–95 (2010). Article ADS MATH Google Scholar * Tejedor, L., Izquierdo, A., Sein, D. V. & Kagan, B. A. Tides and tidal energetics of the
Strait of Gibraltar: A modelling approach. _Tectonophysics_ 294, 333–347 (1998). Article ADS MATH Google Scholar * Harzallah, A., Alioua, M. & Li, L. Mass exchange at the Strait of
Gibraltar in response to tidal and lower frequency forcing as simulated by a Mediterranean Sea model. _Tellus A_ 66, 1–21 (2014). Article Google Scholar * Tesch, F.-W., Niermann, U. &
Plaga, A. Differences in development stage and stock density of larval _Anguilla anguilla_ off the west coast of Europe. _Vie Milieu_ 36, 255–260 (1986). MATH Google Scholar * Marohn, L.
et al. Long-term reduction of late-stage European eel larval abundance at the continental slope reflects glass eel recruitment decline. _ICES J. Mar. Sci._ _Under review_ * Melià, P. et al.
Integrating field data into individual-based models of the migration of European eel larvae. _Mar. Ecol. Prog. Ser._ 487, 135–149 (2013). Article ADS MATH Google Scholar * Wuenschel, M.
J. & Able, K. W. Swimming ability of eels (_Anguilla rostrata_ and _Conger oceanicus_) at estuarine ingress: Contrasting patterns of cross-shelf transport?. _Mar. Biol._ 154, 775–786
(2008). Article Google Scholar * Gonzalez, N. et al. Understanding tidal mixing at the Strait of Gibraltar: A high-resolution model approach. _Prog. Oceanogr._ 212, 102980 (2023). Article
MATH Google Scholar * Izquierdo, A. et al. Modelling in the Strait of Gibraltar: From operational oceanography to scale interactions. _Fundam. Prikl. Gidrofiz._ 9, 15–24 (2016). MATH
Google Scholar * Cresci, A. et al. Glass eels (_Anguilla anguilla_) have a magnetic compass linked to the tidal cycle. _Sci. Adv._ 3, 1–9 (2017). Article MATH Google Scholar * Durif, C.
M. F. et al. Whether European eel leptocephali use the Earth’s magnetic field to guide their migration remains an open question. _Curr. Biol._ 27, R998–R1000 (2017). Article CAS PubMed
Google Scholar * Durif, C. M. F. et al. A unifying hypothesis for the spawning migrations of temperate anguillid eels. _Fish Fish._ 23, 358–375 (2022). Article MATH Google Scholar *
Walsh, P. S., Metzger, D. A. & Higuchi, R. Chelex 100 as a medium for simple extraction of DNA for PCR-based typing from forensic material. _Biotechniques_ 10, 506–513 (1991). CAS
PubMed MATH Google Scholar * Altschul, S. F., Gish, W., Miller, W., Myers, E. W. & Lipman, D. J. Basic local alignment search tool. _J. Mol. Biol._ 215, 403–410 (1990). Article CAS
PubMed MATH Google Scholar * Hanel, R. et al. ADCP current measurements (38 kHz) during RV METEOR cruise M185. PANGAEA https://doi.org/10.1594/PANGAEA.956604 (2023). * Lorente, P. et al.
Accuracy assessment of high-frequency radar current measurements in the Strait of Gibraltar. _J. Oper. Oceanogr._ 7, 59–73 (2014). MATH Google Scholar * Lorente, P. et al. Long-term
monitoring of the Atlantic Jet through the Strait of Gibraltar with HF radar observations. _J. Mar. Sci. Eng._ 7, 3 (2019). Article ADS MATH Google Scholar * Lorente, P. et al. Coastal
high-frequency radars in the Mediterranean—Part 1: Status of operations and a framework for future development. _Ocean Sci._ 18, 761–795 (2022). Article ADS MATH Google Scholar Download
references ACKNOWLEDGEMENTS The scientific crew of M185 gratefully acknowledges the very friendly and most effective cooperation with Captain Detlef Korte and the entire METEOR crew. We also
thank Benedikt Merk, Silvia Blum, Peter Müller, Greta Voss, Kira Kremer and Sebastian Weis for their help on deck and in the laboratory. Many thanks also to the two reviewers in the
peer-review process, who have greatly helped to improve this publication with their pertinent and valuable comments and suggestions. Further, we appreciate the valuable support of the German
Research Fleet Coordination Centre (Leitstelle Deutsche Forschungsschiffe) at the University of Hamburg. FUNDING Open Access funding enabled and organized by Projekt DEAL. The R/V METEOR
expedition M185 was funded by the Deutsche Forschungsgemeinschaft—DFG. J.D.P, L.M. and M.F. are co-funded by the European Commission’s Data Collection Framework (DCF). AUTHOR INFORMATION
AUTHORS AND AFFILIATIONS * Thünen Institute of Fisheries Ecology, Federal Research Institute for Rural Areas, Forestry and Fisheries, 27572, Bremerhaven, Germany Marko Freese, Lasse Marohn,
Jan-Dag Pohlmann, Klaus Wysujack, Tina Blancke & Reinhold Hanel * AZTI, Marine Research, Basque Research and Technology Alliance (BRTA), Herrera Kaia, Portualdea z/g, 20110, Pasaia,
Spain Luis Ferrer Authors * Marko Freese View author publications You can also search for this author inPubMed Google Scholar * Lasse Marohn View author publications You can also search for
this author inPubMed Google Scholar * Luis Ferrer View author publications You can also search for this author inPubMed Google Scholar * Jan-Dag Pohlmann View author publications You can
also search for this author inPubMed Google Scholar * Klaus Wysujack View author publications You can also search for this author inPubMed Google Scholar * Tina Blancke View author
publications You can also search for this author inPubMed Google Scholar * Reinhold Hanel View author publications You can also search for this author inPubMed Google Scholar CONTRIBUTIONS
M.F. and L.M. conceptualized the study. M.F., L.M., L.F., T.B., J.D.P., K.W., and R.H. developed and used the methodology. M.F., L.M., L.F., and T.B. performed the formal analysis. M.F.,
L.M., K.W., L.F., and R.H. provided resources. M.F., L.F., T.B., and L.M. curated the data. M.F., L.F., J.D.P., and T.B. prepared the visualizations. M.F. wrote the original draft of the
manuscript. All authors reviewed and edited the manuscript. CORRESPONDING AUTHOR Correspondence to Marko Freese. ETHICS DECLARATIONS COMPETING INTERESTS The authors declare no competing
interests. ETHICAL STATEMENT Animal ethical approval was not applicable for sampling in this study. ADDITIONAL INFORMATION PUBLISHER’S NOTE Springer Nature remains neutral with regard to
jurisdictional claims in published maps and institutional affiliations. SUPPLEMENTARY INFORMATION SUPPLEMENTARY INFORMATION. RIGHTS AND PERMISSIONS OPEN ACCESS This article is licensed under
a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate
credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article
are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and
your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this
licence, visit http://creativecommons.org/licenses/by/4.0/. Reprints and permissions ABOUT THIS ARTICLE CITE THIS ARTICLE Freese, M., Marohn, L., Ferrer, L. _et al._ Details on the transport
of European eel larvae through the Strait of Gibraltar into the Mediterranean Sea. _Sci Rep_ 15, 1006 (2025). https://doi.org/10.1038/s41598-024-82929-z Download citation * Received: 13
September 2024 * Accepted: 10 December 2024 * Published: 06 January 2025 * DOI: https://doi.org/10.1038/s41598-024-82929-z SHARE THIS ARTICLE Anyone you share the following link with will be
able to read this content: Get shareable link Sorry, a shareable link is not currently available for this article. Copy to clipboard Provided by the Springer Nature SharedIt content-sharing
initiative KEYWORDS * _Anguilla anguilla_ * Recruitment * Leptocephalus * Tidal cycles * Internal waves